

Hiểu các sự kiện vận động: một nghiên cứu sinh lý thần kinh



Tóm tắt. Các neuron phần đuôi vỏ não tiền vận động của khỉ phóng điện trong các chuyển động tay hướng đến mục tiêu như nắm bắt, giữ và xé rách. Ở đây, chúng tôi báo cáo rằng, nhiều trong số các neuron này cũng trở nên hoạt động khi con khỉ quan sát các chuyển động biệt định, có ý nghĩa, của tay, trong các thực hiện thực nghiệm. Các chuyển động có hiệu quả trong thực nghiệm bao gồm đặt hoặc lấy một miếng thức ăn trên bàn, nhận lấy thức ăn từ tay của người làm thí nghiệm và thao tác với các đồ vật. Luôn có một mối liên hệ rõ ràng giữa chuyển động hiệu quả được quan sát và chuyển động được thực hiện, bởi chính con khỉ, và thông thường, chỉ những chuyển động thực nghiệm giống hệt với những chuyển động thực hiện bởi cùng một neuron nhất định mới có thể kích hoạt nó. Những phát hiện này chỉ ra rằng các neuron tiền vận động có thể bị truy xuất trong các chuyển động không chỉ trên cơ sở các đặc điểm kích thích, như đã mô tả trước đây, mà còn trên cơ sở ý nghĩa của các hành động quan sát được.
Tổng quan
Người ta đã xác định rõ, rằng ở khỉ, có một vùng tương ứng với chuyển động đầu xa cánh tay trong vỏ não tiền vận động dưới (Rizzolatti và cộng sự, 1981; Kurata và Tanji 1986; Rizzolatti và cộng sự, 1988; xem thêm Matsumura và Kubota 1979; Muakkassa và Strick 1979). Vùng này nằm ở phía đuôi gần rãnh cung/ arcuate sulcus, và cùng tính mô hoá sinh mở rộng với khu vực F5 (Matelli và cộng sự, 1985). Một trong những đặc điểm phân biệt của neuron F5 là chúng trở nên hoạt động trong các chuyển động tay hướng đến mục tiêu cụ thể, chẳng hạn như nắm, giữ và xé rách. Nhiều trong số chúng dành riêng cho các loại cầm nắm tay khác nhau. Một số phóng điện trong quá trình cầm bằng ngón trỏ và ngón cái, một số trong quá trình cầm nắm bằng ngón tay và một số khác nữa trong quá trình cầm nắm bằng toàn bộ bàn tay. Hơn nữa, một tỷ lệ neuron F5 được kích hoạt bởi các kích thích thị giác, đòi hỏi một kiểu cầm nắm riêng biệt (Rizzolatti và cộng sự, 1988). Mục đích ban đầu của chúng tôi trong các thực nghiệm hiện tại là nghiên cứu hoạt động của các neuron F5 trong một tình huống hành vi, trong đó chúng tôi có thể tách các phản ứng liên quan đến kích thích khỏi hoạt động liên quan đến chuyển động. Với mục đích này, một con khỉ macaque đã được huấn luyện để lấy các vật thể có kích thước và hình dạng khác nhau từ hộp thử nghiệm với độ trễ thay đổi sau khi trình bày kích thích. Sau các thí nghiệm được ghi chép ban đầu, chúng tôi tình cờ quan sát thấy rằng một số hành động của người thí nghiệm, chẳng hạn như nhặt thức ăn hoặc đặt nó vào trong hộp thử nghiệm, đã kích hoạt một tỷ lệ tương đối lớn các neuron F5 mà không có bất kỳ chuyển động công khai nào của khỉ. Mục đích của giao tiếp này là để mô tả một số đặc điểm thiết yếu mới đáng ngạc nhiên này của lớp neuron tiền vận động.
Phương pháp
Các thực nghiệm được thực hiện trên một con khỉ Nemestrina Macaca. Chuẩn bị động vật và quy trình thí nghiệm cơ bản giống như trong các thí nghiệm trước đây của chúng tôi (để biết chi tiết, xem Rizzolatti và cộng sự, 1988, 1990). Trong quá trình ghi, con khỉ ngồi trên một chiếc ghế thí nghiệm với đầu cố định. Các neuron đơn lẻ được ghi lại từ dưới khu vực 6 (khu vực F5) bằng cách sử dụng các điện cực vi mô vonfram (trở kháng 0,5-2 MΩ được đo ở tần số 1 kHz). Vi điện cực được sử dụng để ghi cũng được sử dụng để kích thích vi mô. Kích thích được thực hiện trong mỗi lần thâm nhập sâu 500 µm bằng cách tạo các chuỗi xung cực âm được tạo ra bởi một bộ kích thích dòng điện không đổi (thời lượng tập luyện 50 ms, thời lượng xung 2 ms, tần số 250 Hz, cường độ dòng điện 3-40 µA).
Các neuron lần đầu tiên được thử nghiệm không chính thức bằng cách trình bày các vật thể có kích thước và hình dạng khác nhau ở các vị trí không gian khác nhau (để biết chi tiết về quy trình thử nghiệm, xem Rizzolatti và cộng sự, 1988). Khi rõ ràng rằng một neuron kích hoạt trong quá trình chuyển động tay của khỉ, sự phóng điện của nó đã được nghiên cứu trong một tình huống được kiểm soát hành vi. Một hộp thử nghiệm (45 cm x 30 cm x 35 cm) được đặt trước con khỉ, ngang vai của nó. Cửa trước hộp được tạo thành bởi một chiếc gương một chiều. Các chất rắn hình học có kích thước và hình dạng khác nhau được đặt bên trong hộp. Ánh sáng phòng được điều chỉnh để ngăn con vật nhìn thấy bên trong hộp. Con khỉ bắt đầu các thử nghiệm bằng cách nhấn một công tắc, được tạo thành bởi hai tấm mỏng, bằng ngón cái và ngón trỏ. Nhấn công tắc sẽ thắp sáng hộp và làm cho đối tượng có thể nhìn thấy. Sau một khoảng thời gian trễ 1,2-1,5 giây, cánh cửa mở ra và con khỉ được phép với lấy vật thể. Con vật được thưởng một miếng thức ăn nằm dưới vật thể. Nếu con khỉ nhả công tắc trước khi cửa mở ra, phiên sẽ bị hủy bỏ. Chuyển động của cánh tay và bàn tay được ghi lại bằng cách sử dụng hệ thống ghi lại chuyển động trên máy tính (hệ thống ELITE, Ferrigno và Pedotti, 1985). Hệ thống này, như được sử dụng trong thí nghiệm hiện tại, bao gồm hai camera truyền hình hồng ngoại và một bộ xử lý xây dựng các hình ảnh video trong thời gian thực và tái tạo các vị trí ba chiều của các điểm đánh dấu phản xạ hồng ngoại. Các điểm đánh dấu được sử dụng để tái tạo chuyển động của bàn tay và cánh tay được đặt trên các đốt đầu tiên của ngón trỏ và ngón cái và trên mỏm trâm quay. Hoạt động thần kinh, ghi động học và dấu thời gian sự kiện (bật đèn, mở cửa) được hệ thống ELITE thu thập và ghi lại trên đĩa từ để phân tích ngoại tuyến.
Sau khi phát hiện ra rằng hành động trong một số thực nghiệm có thể kích hoạt các neuron F5, tất cả các neuron được ghi lại đã được kiểm tra bằng cách thực hiện một loạt các hành động vận động trước mặt động vật. Những hành động này có liên quan đến việc nắm lấy thức ăn (ví dụ: đưa thức ăn cho khỉ, đặt nó trên một bề mặt, lấy nó, đưa nó cho người thí nghiệm thứ hai, lấy nó khỏi anh ta), thao tác với thức ăn và các đồ vật khác (bẻ gãy, xé rách, gấp lại), hoặc là cử chỉ có hoặc không có nội dung cảm xúc (đe dọa, nâng tay, vẫy tay). Ngoài ra, các hành động sau đây cũng được thực hiện để đối chứng liệu các neuron được ghi lại có mã hóa cụ thể các mối quan hệ giữa bàn tay và đối tượng hay không: chuyển động của bàn tay mà không có thức ăn, việc chuẩn bị thức ăn bằng một công cụ (ví dụ: kẹp, kìm), các chuyển động kết hợp đồng thời của thức ăn và bàn tay tách biệt bởi khoảng không gian. Tất cả các hành động của người thí nghiệm được lặp lại ở bên phải và bên trái của khỉ ở các khoảng cách khác nhau (50 cm, 1 và 2 m). Hành vi của động vật và hành động của người thí nghiệm trong quá trình thử nghiệm được ghi lại trên một bản ghi video; Hoạt động thần kinh được ghi lại đồng thời trên đường đồ thị song song để tương quan hành vi của con khỉ hoặc hành động của người thí nghiệm với đơn vị phóng điện.
Cơ cánh tay và cẳng tay, bao gồm cơ gấp và cơ duỗi của các ngón tay, được ghi lại đơn cực bằng các điện cực bằng thép không gỉ. Các bản ghi điện cơ được thực hiện trong các phiên đặc biệt, trong đó các quy trình thử nghiệm giống hệt với các quy trình trong đó các neuron được ghi lại.
Kết quả
Hoạt động của hai neuron trong quá trình khỉ nắm bắt/ grasping các vật thể nhỏ và lớn được minh họa trong Hình 1. Đơn vị 483 truyền dẫn mạnh trong quá trình cầm nắm chính xác (bảng phía trên), trong khi nó chỉ được kích hoạt yếu trong quá trình xử lý bằng toàn bộ bàn tay (bảng phía dưới). Đơn vị 481 không cho thấy bất kỳ tính đặc hiệu nào đối với kiểu nắm bắt. Sự phóng điện của nó, vốn tự nhiên cao, bị ức chế mạnh mẽ trong tất cả các kiểu nắm bắt. Hoạt động tự phát của hai neuron không bị ảnh hưởng bởi sự trình bày đối tượng cũng như bởi sự chuẩn bị chuyển động trong thời gian chờ đợi.
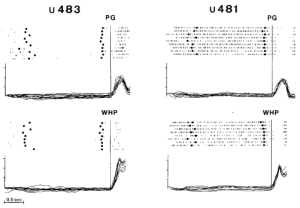 Hình 1. Sự phóng điện của hai neuron tiền vận động trong lúc cầm nắm tay. Hoạt động của neuron (hiển thị ma trận) và các bản ghi động học của hoạt động cầm nắm (môđun của khoảng cách vectơ giữa ngón cái và ngón trỏ) được hiển thị. Các bảng trên: phản ứng của neuron trong quá trình cầm nắm chính xác (PG); các bảng dưới: phản ứng của neuron trong quá trình cầm nắm toàn bộ bàn tay (WHP). Các vật thể là một quả cầu nhỏ (đường kính 0,4 cm) trong trường hợp PG và một quả cầu lớn (đường kính 4 cm) cho đơn vị 483 và một hình trụ lớn (đường kính 2 cm) cho đơn vị 481 trong trường hợp WHP. Sự phóng điện của neuron và các bản ghi động học được căn chỉnh với thời điểm bắt đầu mở tay (các đường thẳng đứng liên tục). Chấm đầu tiên được tô đậm của mỗi ma trận đánh dấu sự trình bày của vật thể; chấm thứ hai được tô đậm đánh dấu thời điểm vật thể được cầm nắm. Mỗi phân đoạn biểu thị 20 ms. Sắp xếp theo thứ tự: mỗi phân đoạn biểu thị 0,7 cm
Hình 1. Sự phóng điện của hai neuron tiền vận động trong lúc cầm nắm tay. Hoạt động của neuron (hiển thị ma trận) và các bản ghi động học của hoạt động cầm nắm (môđun của khoảng cách vectơ giữa ngón cái và ngón trỏ) được hiển thị. Các bảng trên: phản ứng của neuron trong quá trình cầm nắm chính xác (PG); các bảng dưới: phản ứng của neuron trong quá trình cầm nắm toàn bộ bàn tay (WHP). Các vật thể là một quả cầu nhỏ (đường kính 0,4 cm) trong trường hợp PG và một quả cầu lớn (đường kính 4 cm) cho đơn vị 483 và một hình trụ lớn (đường kính 2 cm) cho đơn vị 481 trong trường hợp WHP. Sự phóng điện của neuron và các bản ghi động học được căn chỉnh với thời điểm bắt đầu mở tay (các đường thẳng đứng liên tục). Chấm đầu tiên được tô đậm của mỗi ma trận đánh dấu sự trình bày của vật thể; chấm thứ hai được tô đậm đánh dấu thời điểm vật thể được cầm nắm. Mỗi phân đoạn biểu thị 20 ms. Sắp xếp theo thứ tự: mỗi phân đoạn biểu thị 0,7 cm
Hình 2 và 3 cho thấy phản ứng của các đơn vị 483 và 481 trong khi con khỉ quan sát các chuyển động nắm bắt được thực hiện bởi người làm thực nghiệm. Mối tương quan giữa các chuyển động và hoạt động thần kinh được thực hiện trong trường hợp này được ghi trong các bản ghi video. Biến đổi trong phóng điện thần kinh được xác định rõ ràng trong quá trình quan sát nắm bắt (Hình 2A, 3A, B) cũng như trong quá trình thực hiện nắm bắt (Hình 2 B, 3 C). Mối quan tâm đặc biệt là Hình 3 B. Ở đây các hồ sơ cho thấy hoạt động của neuron không bị gián đoạn từ thời điểm người thực nghiệm nhặt thức ăn (mũi tên đầu tiên) cho đến khi khỉ nắm lấy nó (mũi tên thứ hai). Cả việc nắm bắt được quan sát và nắm bắt được thực hiện đều ức chế sự phóng điện của neuron. Ngược lại, cả chuyển động của bàn tay của người thực nghiệm về phía thức ăn (ghi trước mũi tên đầu tiên) cũng như việc trình bày thức ăn cho khỉ (ghi trước mũi tên thứ hai) đều không ảnh hưởng đến việc phóng điện thần kinh. Lưu ý rằng sau khi người thực nghiệm hành động nắm bắt, lượng phóng điện đã trở lại tốc độ cao mặc dù thực tế là sau khi nắm bắt, thức ăn được di chuyển về phía con vật và do đó, sự quan tâm và chuẩn bị của khỉ để thực hiện các chuyển động cánh tay và tay tăng lên.
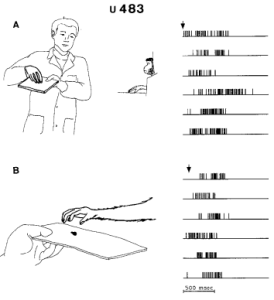 Hình 2A, B. Ví dụ về một đơn vị phóng điện chọn lọc trong khi khỉ chuyển động nắm bắt và trong khi khỉ quan sát các chuyển động nắm bắt do người thực nghiệm thực hiện. A Người thực nghiệm nắm bắt thức ăn; B khỉ nắm bắt thức ăn. Các mũi tên chỉ ra thời điểm bắt đầu nắm bắt (xấp xỉ). Quy trình chuẩn cho đơn vị này (483) được thể hiện trong Hình 1, bên trái
Hình 2A, B. Ví dụ về một đơn vị phóng điện chọn lọc trong khi khỉ chuyển động nắm bắt và trong khi khỉ quan sát các chuyển động nắm bắt do người thực nghiệm thực hiện. A Người thực nghiệm nắm bắt thức ăn; B khỉ nắm bắt thức ăn. Các mũi tên chỉ ra thời điểm bắt đầu nắm bắt (xấp xỉ). Quy trình chuẩn cho đơn vị này (483) được thể hiện trong Hình 1, bên trái
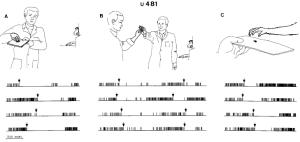
Hình 3A-C. Ví dụ về một đơn vị phóng điện chọn lọc trong lúc khỉ nắm bắt chuyển động và trong lúc khỉ quan sát chuyển động nắm bắt của người thực nghiệm. Các tình huống trong A và C tương tự như trong Hình 2A và B. Trong B Người thực nghiệm lấy một miếng thức ăn từ một người thực nghiệm khác (mũi tên đầu tiên) và đưa nó về phía con khỉ. Mũi tên thứ hai chỉ ra thời điểm con khỉ nắm lấy thức ăn từ tay người thực nghiệm. Quy chuẩn thực hiện của đơn vị này (481) được thể hiện trong Hình 1, bên phải
Tổng cộng có 184 neuron F5 đã được nghiên cứu. Đồng dạng với những phát hiện trước đây, hầu hết tất cả các neuron đều hoạt động trong các loại chuyển động xa khác nhau (cắm, giữ, xé). Kích thích vi mô được thực hiện tương ứng với các vị trí mà từ đó các neuron được ghi lại đã gợi ra chuyển động của bàn tay và ngón tay. Tám mươi bảy neuron phản ứng với các kích thích thị giác. Bốn mươi tám tế bào trong số đó phản ứng với các kích thích có ý nghĩa đơn giản (ví dụ: thức ăn) như đã mô tả trước đây (Rizzolatti và cộng sự, 1988), trong khi 39 tế bào có các đặc tính thị giác phức tạp tương tự như các đơn vị 481 và 483. Các neuron có đặc tính thị giác phức tạp được chia thành bốn loại lớn. Loại đầu tiên (n = 12) bao gồm các đơn vị trong đó quan sát hành động hiệu quả và thực hiện hành động được hiệu quả tương ứng. Hành động được mã hóa có thể là nắm bằng miệng, nắm bằng tay (thường xuyên được thể hiện nhất), xoay đối tượng hoặc thao tác đối tượng. Loại thứ hai (n = 6) được hình thành bởi các neuron, trong đó hành động quan sát hiệu quả là hành động hiệu quả do chính con khỉ thực hiện (ví dụ: nắm bắt) cộng với các hành động khác tương tự như hành động được thực hiện (ví dụ: đặt một vật trên bàn). Loại thứ ba (n = 11) bao gồm các neuron trong đó các hành động quan sát hiệu quả có liên quan hợp lý đến các hành động được thực hiện hiệu quả và có thể được coi là sự chuẩn bị cho chúng. Ví dụ, hành động quan sát hiệu quả là đặt một vật trên bàn, trong khi hành động được thực hiện hiệu quả là đưa thức ăn vào miệng hoặc nắm lấy đồ vật. Lớp cuối cùng (n = 10) được hình thành bởi các neuron phản ứng với các hành động quan sát được tương tự như những hành động được mô tả ở trên, nhưng không có hoạt động tương quan với chuyển động của động vật. Các chuyển động tích cực của động vật mà không có sự kiểm soát thị giác có hiệu quả trong việc kích hoạt các neron của ba lớp đầu tiên. Tuy nhiên, không phải tất cả các neron đều được thử nghiệm trong trường hợp không có kiểm soát thị giác.
Thử nghiệm đối chứng về tính đặc hiệu của mối quan hệ giữa bàn tay-đối tượng (xem Phương pháp) cho thấy các chuyển động của bàn tay của người thực nghiệm hoặc các chuyển động kết hợp của bàn tay và vật thể tách biệt về mặt không gian với nhau không hiệu quả trong việc kích hoạt các neuron của tất cả các lớp. Tương tự, nắm thức ăn bằng kẹp, kìm hoặc các dụng cụ khác không ảnh hưởng đến phóng điện thần kinh. Các chuyển động đe dọa cũng không tạo hiệu quả.
Các chuyển động thích hợp của người thực nghiệm kích hoạt các neron cho dù được thực hiện trong không gian gần cận của khỉ hay xa nó. Tuy nhiên, các phản ứng thường mạnh hơn khi các kích thích quan sát được ở gần với con vật. Đối với một số đơn vị, phía kích thích ảnh hưởng đến cường độ phản ứng.
Để xác minh liệu sự kích hoạt neuron trong các hành động quan sát có thể liên quan đến chuyển động của động vật mà sự quan sát là không chú ý hay không, các chuyển động bên ngoài của cánh tay và bàn tay của con khỉ đã được ghi lại điện cơ trong các hành động của các nhà thí nghiệm. Các chuyển động đầu xa được theo dõi vì những chuyển động này tương ứng trên F5 và vì phân tích định lượng của các neuron phóng điện liên quan đến hành động trong các thực nghiệm cho thấy mối tương quan rõ ràng, trong hầu hết các trường hợp, giữa các phóng điện và chuyển động đầu xa. Những thay đổi của hoạt động điện cơ liên quan đến hành động quan sát được không bao giờ được tìm thấy.
Thảo luận
Một trong những chức năng cơ bản của vỏ não tiền vận động là truy xuất các hành động vận động thích hợp để đáp ứng với các kích thích cảm giác. Bằng chứng đã được cung cấp cho thấy việc truy xuất hành động có thể xảy ra để đáp ứng với các mẫu hai chiều (Mitz và cộng sự, 1991), màu sắc (Halsband và Passingham 1985; Passingham 1988), và kích thước và hình dạng của các vật thể ba chiều (Petrides 1982; Rizzolatti và cộng sự, 1988; Gentilucci và Rizzolatti 1990). Dữ liệu hiện tại chỉ ra rằng ngoài những yếu tố vật lý này, việc truy xuất cũng có thể xảy ra để đáp ứng ý nghĩa của các cử chỉ do các cá nhân khác thực hiện. Nếu người ta xem xét các tương tác xã hội phong phú trong một nhóm khỉ, sự hiểu biết của một con khỉ về các hành động được thực hiện bởi những con khỉ khác phải là một yếu tố rất quan trọng trong việc xác định lựa chọn hành động. Do đó, khả năng của các neuron tiền vận động phía dưới để lựa chọn các hành động theo ý nghĩa cử chỉ phù hợp với khuôn khổ khái niệm của lý thuyết hiện tại về các chức năng của vỏ não tiền vận động và mở rộng nó để bao gồm lựa chọn chuyển động liên quan đến các mối quan hệ giữa các cá nhân.
Khám phá vỏ não tiền vận động với các neuron nhạy cảm với ý nghĩa của hành động là điều bất ngờ. Tuy nhiên, sự hiện diện của các neuron này phù hợp với những phát hiện trước đó. Perrett và các đồng nghiệp (1989, 1990) đã mô tả các neuron trong thùy thái dương mà có phản ứng phụ thuộc vào mối quan hệ giữa chuyển động của bàn tay và vật thể. Các đặc điểm của các neuron này rất giống với các đặc điểm của nhóm cuối mà chúng tôi đã nêu, thuần túy liên quan đến thị giác. Do đó, có vẻ như các tương tác giữa bàn tay và đối tượng thì thuộc về các loại kích thích phức tạp, giống như khuôn mặt (Gross và cộng sự, 1972; Rolls 1984; Perrett và cộng sự 1984, 1987; Desimone 1991), được mã hóa rõ ràng bởi các neuron riêng lẻ. Tầm quan trọng của hành vi lựa chọn nhanh các chuyển động thích hợp theo chuyển động của các cá nhân khác có lẽ đã ủng hộ loại mã hóa này, cho phép nhận biết nhanh các kích thích.
Một phát hiện rất thú vị khác trong các thí nghiệm của chúng tôi là quan sát thấy một số neuron chỉ được kích hoạt bởi chuyển động của các nhà thực nghiệm trùng khớp với những neuron kích hoạt neuron khi được thực hiện bởi khỉ. Có các tài liệu báo cáo rằng bệnh nhân mắc triệu chứng mất ý tưởng vận động/ ideomotor apraxia có thể gặp khó khăn (Heilman và cộng sự, 1982) trong việc nhận ra ý nghĩa của các cử chỉ do người khác thực hiện. Những phát hiện này được coi là bằng chứng cho thấy tri giác cử chỉ và vận động/praxia của các chi chia sẻ cùng một mạch vỏ não (Rothi và cộng sự, 1991). Tương tự, phát hiện rằng các mẫu âm thanh của lời nói tổng hợp phải được sửa đổi nếu một nhận thức ngữ âm bất biến được tạo ra trong các bối cảnh khác nhau đã khiến một số tác giả đề xuất rằng các đối tượng của tri giác lời nói không được tìm thấy trong âm thanh, mà trong các cử chỉ ngữ âm/ phonetic gestures của người nói, được thể hiện trong não dưới dạng các lệnh vận động bất biến (xem Liberman và Mattingly, 1985). Mặc dù các quan sát của chúng tôi không có nghĩa là chứng minh các lý thuyết vận động của tri giác, tuy nhiên chúng chỉ ra rằng trong các khu vực vỏ não tiền vận động có các neuron được ban tặng các đặc tính mà các lý thuyết đó yêu cầu. Thật thú vị khi lưu ý rằng vị trí giải phẫu của khu vực dưới vùng 6, và đặc biệt là F5, tương ứng phần lớn với vị trí giải phẫu của vùng Broca trong não người (Freund, 1991).
Một thực nghiệm khác được nhóm nghiên cứu thực hiện sau đó (1996) xem xét việc ghi nhớ hành động. Đặc biệt nhấn mạnh việc tri nhận và ghi nhớ chuyển động vùng miệng (cơ sở của tri nhận ngôn ngữ).



- Bs Đỗ Thị Thúy Anh dịch và soạn