

Học tập kiểu Hebb và dự đoán của neuron gương với hành động, cảm giác và cảm xúc



TÓM TẮT
Tính khả biến phụ thuộc-thời gian-nhọn sóng được coi là cơ sở sinh lý thần kinh của việc Học tập kiểu Hebb và đã được chứng minh là nhạy cảm với cả tính ngẫu nhiên và sự tiếp giáp giữa hoạt động trước và sau synap. Ở đây, chúng tôi sẽ xem xét cách áp dụng quy tắc học tập kiểu Hebb này cho một hệ thống các neuron được liên hiệp với nhau khi có sự tái biểu hiện trực tiếp hoặc gián tiếp (ví dụ: nhìn/nghe thấy hành động của chính mình) tính dự đoán sự xuất hiện của các neuron gương với các đặc tính có thể dự đoán. Trong khuôn khổ này, chúng tôi phân tích cách các neuron gương trở thành một hệ thống có động lực, thực hiện các suy luận tích cực về hành động của người khác và cho phép tham gia các hành động chung bất chấp sự chậm trễ về cảm biến vận động. Chúng tôi khám phá cách hệ thống này thực hiện việc phóng chiếu bản thân lên người khác, với những thiên kiến kiến bản thân làm trung tâm để góp phần đọc-suy nghĩ. Cuối cùng, chúng tôi lập luận rằng việc học tập kiểu Hebb dự đoán các neuron giống như tấm gương về cảm giác và cảm xúc, đồng thời xem xét bằng chứng về sự hiện diện của những hoạt động đại biểu/vicarious như vậy bên ngoài hệ thống vận động.
GIỚI THIỆU
Việc phát hiện ra các neuron gương cung cấp bằng chứng khoa học thần kinh cho cái mà chúng ta gọi là kích hoạt tương ứng/ representation: cơ sở vật chất thần kinh cho hành động của chính chúng ta được kích hoạt tương ứng trong lúc chứng kiến hành động của người khác thông qua thị giác [1–4] hoặc âm thanh [3,4]. Hai mươi năm sau khi được phát hiện, chức năng của neuron gương vẫn còn được tranh luận sôi nổi [5–9]. Ở đây, chúng tôi không giải quyết câu hỏi về chức năng của chúng mà chỉ khám phá cách chúng có thể phát triển. Khỉ có neuron gương phản ứng với âm thanh và hình ảnh khi vò nát một túi nhựa [3,4] và vỏ não tiền vận động của con người phản ứng với những âm thanh như tiếng rít khi mở lon Coca-Cola [10]. Sự chọn lọc như vậy khó có thể được lập trình trước về mặt di truyền. Ở đây, chúng tôi khám phá góc nhìn cơ chế về cách các neuron gương như vậy có thể xuất hiện trong quá trình phát triển. Chúng tôi xác định những gì khoa học thần kinh hiện đại hiểu được qua cách học tập kiểu Hebb, dựa trên độ khả biến phụ thuộc-thời gian-nhọn sóng (spike-timing-dependent plasticity STDP). Chúng tôi khám phá cách hiểu biết tinh tế về học tập kiểu Hebb này, giúp chúng tôi hiểu cách các neuron gương xuất hiện và gợi ý cách các neuron gương trở thành một hình thức đọc suy nghĩ mang tính dự đoán hoạt động. Cuối cùng, chúng tôi lập luận rằng các kích hoạt tương ứng cũng xảy ra ở vỏ não cảm xúc và cảm giác thân thể, đồng thời các quy tắc học tập tương tự kiểu Hebb có thể giải thích sự xuất hiện của các neuron giống như tấm gương ở các vùng não này.
HỌC TẬP KIỂU HEBB Ý NGHĨA LÀ GÌ
(a) Về mặt lịch sử
Thuật ngữ ‘Học tập kiểu Hebb’ bắt nguồn từ công trình của Donald Hebb [11], người đã đề xuất một giải thích sinh lý thần kinh về học tập và trí nhớ dựa trên một nguyên tắc đơn giản: “Khi một sợi trục của tế bào A ở khoảng cách gần đến mức đủ để kích thích tế bào B và lặp đi lặp lại hoặc liên tục, thì sẽ truyền dẫn các kích thích đó, một số quá trình tăng trưởng hoặc thay đổi trao đổi chất diễn ra ở một hoặc cả hai tế bào sao cho hiệu quả của A, với tư cách là một trong các tế bào kích hoạt B, được tăng lên” (tr. 62). Việc đọc kỹ quy luật của Hebb sẽ cho thấy sự hiểu biết của ông về tầm quan trọng của quan hệ nhân quả và tính nhất quán. Ông viết, không phải rằng hai neuron cần hoạt động cùng nhau để tăng hiệu quả kết nối của chúng, mà là một neuron cần liên tục (nhất quán) tham gia vào việc kích hoạt (quan hệ nhân quả) kia. Carla Shatz (nhưng không phải bản thân Hebb) đã diễn giải nguyên tắc của mình bằng một vần điệu: ‘cái gì cháy cùng nhau, nối với nhau’ [12, tr. 64]. Mặc dù mang tính ghi nhớ nhưng bản tóm tắt này có nguy cơ che khuất tầm quan trọng của quan hệ nhân quả trong công việc thực tế của Hebb: nếu hai neuron cùng hoạt động theo đúng nghĩa đen, tức là cùng một lúc, việc kích hoạt của một neuron này không thể gây ra hoạt động của tế bào kia. Ưu tiên tạm thời, chứ không phải tính đồng thời, là dấu hiệu của quan hệ nhân quả [13] và sẽ chỉ ra rằng ‘cái này tham gia vào việc kích hoạt cái kia’. Do đó, cách diễn giải này hẳn được đọc như chút muối bỏ bể.
(b) Hiểu biết về sinh lý thần kinh
Vào những năm 1990, các nhà sinh lý học thần kinh đã đặt nền móng cho sự hiểu biết sinh lý thần kinh hiện đại của chúng ta về việc Học tập kiểu Hebb dựa trên STDP [14–16]. Các thí nghiệm trong đó hai neuron được kết nối được kích thích với sự khởi phát không đồng bộ của các kích thích khác nhau đã chứng minh một cửa sổ không đối xứng của STDP (hình 1). Khi một synap bị kích thích kết nối với một neuron bị kích thích, nếu neuron tiền synap được kích thích 40 ms hoặc ít hơn trước neuron hậu synap thì synap đó sẽ được tăng cường điện thế. Ngược lại, nếu neuron tiền synap bị kích thích ngay sau neuron hậu synap thì synap sẽ bị ức chế. Nếu hai neuron chỉ đơn giản kích hoạt cùng nhau, thì sự dao động tạm thời không thể tránh khỏi sẽ làm cho neuron tiền synap đôi khi kích hoạt ngay trước và đôi khi ngay sau neuron hậu synap, đồng thời điện thế hóa và ức chế hóa sẽ triệt tiêu lẫn nhau theo thời gian, dẫn đến không có STDP thực đáng kể. Như Hebb đã dự đoán, quan hệ nhân quả do đó là chìa khóa cho tính khả biến của synap.
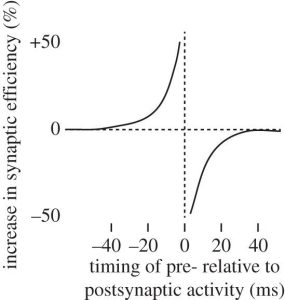 Hình 1. Sự bất đối xứng tạm thời của tính khả biến phụ thuộc-thời gian-nhọn sóng.
Hình 1. Sự bất đối xứng tạm thời của tính khả biến phụ thuộc-thời gian-nhọn sóng.
Các thí nghiệm khác đã cải thiện sự hiểu biết của chúng tôi về tính nhất quán cần thiết để diễn ra tính khả biến của synap. Bauer và cộng sự, [17] đã sử dụng giao thức STDP tiêu chuẩn, với neuron tiền synap được kích thích 5–10 ms trước neuron hậu synap. Áp dụng 10 trong số các kích thích theo cặp này, họ nhận thấy điện thế hóa mạnh mẽ của synap (hình 2a). Việc lặp lại quy trình, nhưng việc trộn lẫn các kích thích không ghép đôi trong đó chỉ có neuron hậu synap được kích thích đã hủy bỏ điện thế mặc dù đã áp dụng chính xác 10 thử nghiệm ghép đôi giống nhau (hình 2b). Điều này chỉ ra rằng tính dự phòng là rất quan trọng đối với STDP: trong hình 2a, hoạt động trước synap dự đoán hoạt động sau synap (p(post|pre) = 1, p(post|no pre) = 0), trong hình 2b, hoạt động trước synap không mang tính thông tin (p(post|pre) = p(post|no pre) = 0,5). Điều này bổ sung thêm những gì Hebb mô tả bằng trực giác là “tham gia lặp đi lặp lại và liên tục” và âm hưởng quy luật về học tập kết hợp [18]. Bauer và cộng sự, [17] sau đó chuyển 10 kích thích không ghép đôi sang sau 10 kích thích ghép đôi và vẫn không tìm thấy điện thế (hình 2c). Tuy nhiên, việc phân phối 10 kích thích chưa được ghép nối, 15 hoặc 50 phút sau các kích thích được ghép nối, không còn hủy STDP nữa (hình 2d). Do đó, các kích thích không ghép đôi sẽ được tích hợp với các kích thích ghép đôi nếu chúng xảy ra trong khoảng thời gian 7 phút để áp dụng 10 thử nghiệm ghép đôi và 10 thử nghiệm không ghép đôi, nhưng sẽ không tích hợp nếu chúng xảy ra muộn hơn nữa. Do đó, STDP phụ thuộc vào cả sự tiếp giáp và tình huống ngẫu nhiên (tùy cơ) và sử dụng khoảng thời gian rất hẹp, khoảng 40 ms, để xác định xem liệu neuron tiền synap có tham gia gây ra một điện thế hoạt động sau synap cụ thể (sự tiếp giáp) hay không và một khoảng thời gian dài hơn nhiều, khoảng 10 phút, để xác định liệu hoạt động trước synap cung cấp thông tin về hoạt động sau synap (tùy cơ). Tuy nhiên, liệu các chi tiết của sự tích hợp tùy cơ này có áp dụng cho tất cả các neuron hay chỉ biệt định cho nhân bên của hạch hạnh nhân hay không vẫn còn phải nghiên cứu.

Hình 2. (a) Áp dụng 10 cặp kích thích trước và sau synap sẽ dẫn đến hiệu ứng đáng kể của synap. (b) Trộn lẫn 10 kích thích sau synap không ghép đôi chỉ làm mất đi điện thế. (c) Áp dụng 10 lần kích thích không ghép đôi sau 10 lần kích thích ghép đôi cũng sẽ hủy bỏ hiệu ứng. (d) Trì hoãn các kích thích không ghép đôi trong 15 hoặc 50 phút sẽ duy trì hiệu lực của 10 thử nghiệm ghép đôi. Kích thích trước synap được biểu thị dưới dạng một đường cong biểu thị điện thế kích thích sau synap đến neuron sau synap, kích thích sau synap là một thanh thẳng đứng để biểu thị điện thế hoạt động. Trích từ Bauer và cộng sự [17]; epsp, điện thế kích thích sau synap
Theo những phát hiện này, ‘Học tập kiểu Hebb’ trong sinh lý thần kinh đương đại đề cập đến sự hiểu biết đang mở rộng nhanh chóng về STDP [15,16], lấy ý tưởng từ công trình của Hebb, và nhấn mạnh tính nhạy cảm của STDP đối với ưu tiên thời gian chặt chẽ (quan hệ nhân quả) và tình huống ngẫu nhiên theo phút. Sự kiên trì của thuật ngữ ‘học tập kiểu Hebb’ để chỉ STDP tôn vinh ký ức của một người đã dự đoán mức độ học tập có thể được giải thích bằng tính khả biến phụ thuộc-thời gian-nhọn sóng như vậy. Ở đây, chúng tôi tuân thủ việc sử dụng cách Học tập kiểu Hebb này. Các ngành khoa học tính toán sử dụng cách hiểu được tinh chỉnh tương tự về việc Học tập kiểu Hebb, điều này cũng phụ thuộc vào tính ngẫu nhiên (http://lcn.epfl.ch/~gerstner/SPNM/node70.html).
(c) Các định nghĩa thay thế
Ngược lại, trong mô tả tâm lý học, một số tác giả vẫn đánh đồng việc Học tập kiểu Hebb với sự liên hệ trong trí nhớ, “những gì cùng truyền dẫn thì kết cùng mạch”. Chúng tôi đặc biệt khám phá định nghĩa thay thế này, được Cooper và cộng sự sử dụng [19], là một ví dụ, bởi vì bài báo đó cố gắng lập luận chống lại việc Học tập kiểu Hebb trong hệ thống neuron gương, và việc hiểu nguồn gốc của sự hiểu lầm là rất quan trọng. Họ viết: Hebb có câu nói nổi tiếng rằng “Các tế bào cùng truyền dẫn thì kết cùng mạch”, và chính thức hơn là, “bất kỳ hai tế bào hoặc hệ thống tế bào nào hoạt động lặp đi lặp lại cùng một lúc sẽ có xu hướng trở nên ‘liên hiệp’, vì hoạt động trong tế bào này tạo thuận cho hoạt động của tế bào kia”. Do đó, quan điểm kiểu Hebb của Keysers và Perrett ngụ ý rằng sự tiếp giáp là đủ cho sự phát triển MNS; rằng nó cũng không phụ thuộc vào sự ngẫu nhiên’.
Chúng tôi nghĩ rằng có một số hiểu lầm trong tuyên bố này. Đầu tiên, bản thân Hebb chưa bao giờ viết “Các tế bào cùng truyền dẫn thì kết cùng mạch”. Cụm từ có tính liên hệ này lần đầu tiên được Carla Shatz [12] giới thiệu trong một bài báo dành cho tạp chí Scientific American nhắm đến công chúng. Thứ hai, điều được trích dẫn gọi là định đề hình thức của Hebb ‘hai tế bào bất kỳ…’, thì không phải vậy. Hebb dùng câu này để tóm tắt những ý tưởng cũ: ông viết ‘Ý tưởng chung là một ý tưởng cũ, rằng hai tế bào bất kỳ…’ [tr. 70]. Cả cụm từ liên hệ được gán sai cho Hebb và tóm tắt các ý tưởng cũ của Hebb đã che phủ yếu tố nhân quả trong định đề thực sự của Hebb ‘Khi một sợi trục của tế bào A ở đủ gần để kích thích một tế bào B và tham gia vào việc kích hoạt nó lặp đi lặp lại hoặc liên tục, một số quá trình tăng trưởng hoặc sự thay đổi trao đổi chất diễn ra trong một hoặc cả hai tế bào sao cho ảnh hưởng của A, với tư cách là một trong các tế bào truyền dẫn sang B, được tăng lên’ ([11], trang 62). Kết quả của những hiểu lầm này là quan niệm của Cooper và những người khác về việc Học tập kiểu Hebb khác với quan điểm của chúng ta: quan điểm của họ tập trung vào tính tiếp giáp, trong khi quan điểm của chúng ta bao gồm quyền ưu tiên theo thời gian, quan hệ nhân quả và tình huống ngẫu nhiên. Bản thân Hebb có thể là người duy nhất biết chính xác liệu ông có thích định nghĩa của chúng tôi hơn định nghĩa của Cooper và cộng sự hay không, nhưng điều quan trọng là phải hiểu sự khác biệt về định nghĩa này để tránh làm những gì chúng tôi tin rằng Cooper và cộng sự đã thực hiện: sử dụng định nghĩa dựa trên sự liền kề của riêng họ và áp dụng nó vào lý thuyết của chúng ta về sự xuất hiện của các neuron gương dựa trên một khái niệm khác về việc Học tập kiểu Hebb. Làm như vậy sẽ dẫn đến sự hiểu lầm về lý thuyết của chúng ta và trong trường hợp này là những tuyên bố không có cơ sở chống lại lý thuyết của chúng ta.
HỌC TẬP KIỂU HEBB VÀ NEURON GƯƠNG: GÓC NHÌN THỜI GIAN-VĨ MÔ
Các neuron gương tồn tại ít nhất ở vùng vỏ não tiền vận động bụng của khỉ (PM; vùng F5, [2–4,20]) và đỉnh sau dưới (vùng PF/PFG, [21]). Các neuron ở hai vùng này được kết nối tương hỗ [22]: PF/PFG gửi thông tin đến PM và PM quay lại PF/PFG. Các neuron trong vùng PF/PFG cũng được kết nối qua lại với các neuron ở rãnh thái dương trên (STS [22,23]), một vùng được biết là phản ứng với nhìn chuyển động của cơ thể, khuôn mặt và âm thanh của hành động [24]. Các vùng não khác cũng chứa các neuron gương [25–27], nhưng để minh họa cách giải thích học tập kiểu Hebb theo sự xuất hiện của các neuron gương, về nguyên tắc có thể giải thích sự xuất hiện của các neuron gương, một hệ thống đơn giản chỉ bao gồm hai vùng não, STS và PM, cùng nhau với các kết nối qua lại từ STS đến PM và từ PM đến STS là đủ. Trong phần này, chúng tôi sẽ áp dụng độ phân giải thời gian tương đối thô khoảng 1 giây cho điểm gần đúng đầu tiên của mô tả Hebb về cách các neuron gương có thể phát sinh. Ở cấp độ mô tả này, việc Học tập kiểu Hebb đưa ra các dự đoán ở cấp độ thần kinh, tương tự như điều mà việc học theo trình tự liên hiệp—một mô hình nhận thức ban đầu được phát triển để mô tả sự xuất hiện của hành vi bắt chước [28]—đưa ra ở cấp độ chức năng. Các mô tả gốc giải thích việc Học tập kiểu Hebb ở độ phân tích tạm thời này là của Keysers & Perrett và Del Giudice và cộng sự [24,29], những người mô tả việc học theo trình tự liên hiệp bao gồm Heyes, Brass & Heyes và Cook và cộng sự [28,30,31]. Trong mục sau, chúng tôi xem xét sau đó một thang thời gian tốt hơn để tiết lộ cách các neuron gương có thể tổ chức một hệ thống có động lực để tạo ra các suy luận tích cực.
(a) Tái-hướng đến như một tín hiệu huấn luyện
Ở trẻ sơ sinh (người) và con non (khỉ), chúng ta biết rất ít về tính chọn lọc của các neuron STS và PM có liên quan cũng như các kết nối của chúng. Theo đó, chúng tôi sẽ giả sử các kết nối hai chiều tương đối ngẫu nhiên giữa các neuron trong STS phản ứng với hình ảnh và âm thanh của các hành động khác nhau, và các neuron trong PM mã hóa để thực hiện các hành động tương tự. Các kết nối này đi qua thùy đỉnh sau (đặc biệt là PF/PFG), nhưng để đơn giản, chúng tôi không đề cập rõ ràng đến bước trung gian này.
Khi một cá nhân thực hiện một hành động tay mới, anh ta sẽ nhìn thấy và nghe thấy chính mình thực hiện hành động đó. Đầu vào cảm giác này xuất phát từ hành động của chính một người được gọi là ‘tái-hướng đến’. Xu hướng chung của những đứa trẻ đang phát triển bình thường là nhìn chằm chằm vào bàn tay của mình đảm bảo rằng sự tái-hướng đến như vậy sẽ xảy ra thường xuyên khi trẻ thực hiện những động tác mới [32]. Kết quả là, hoạt động trong neuron PM kích hoạt một hành động cụ thể và hoạt động trong neuron phản ứng với âm thanh và hình ảnh của hành động cụ thể này trong STS, theo giá trị gần đúng đầu tiên (nhưng xem mục sau), sẽ trùng lặp một cách nhất quán và duy trì về mặt thời gian. Ví dụ, một neuron nắm bắt trong STS sẽ có hoạt động kích hoạt liên tục trùng lặp về thời gian với hoạt động của các neuron nắm bắt PM trong khi cá nhân quan sát thấy chính mình đang nắm bắt. Mặt khác, các neuron STS ném sẽ có hoạt động truyền dẫn liên tục trùng lặp về thời gian với các neuron PM ném trong khi cá nhân quan sát thấy chính mình ném. Ngược lại, việc kích hoạt các neuron nắm bắt STS sẽ không trùng lặp một cách có hệ thống về thời gian với việc kích hoạt các neuron ném PM và ngược lại. Theo đó, sự tái-hướng đến sẽ tạo ra một tình huống trong đó việc kích hoạt các neuron STS và PM cho cùng một hành động sẽ chồng lên một cách có hệ thống hơn so với việc kích hoạt các neuron dành cho hai hành động khác nhau. Có một sự tiếp giáp thô (truyền dẫn vào cùng một thời điểm) và ngẫu nhiên (ví dụ: p(cảnh nắm|thực hiện nắm bắt) > p(cảnh ném|thực hiện nắm bắt)). Ở quy mô thời gian vĩ mô này, các synap kết nối các biểu diễn STS và PM của cùng một hành động phải được tăng cường dựa trên sự hiểu biết về việc Học tập kiểu Hebb đã nêu ở trên, trong khi những synap đại diện cho các hành động khác sẽ bị suy yếu.
(b) Sự Tái-hướng đến sẽ ưu tiên các kết nối phù hợp hơn
Chúng tôi đưa ra giả thuyết rằng, sau nhiều lần tái-hướng đến và Học tập kiểu Hebb, điều đó sẽ khiến các kết nối STS-PM phổ biến phải khớp với nhau (tức là các kết nối tạo biểu diễn của hành động tương tự). Điều này dựa trên giả định mà phần lớn chưa được kiểm chứng, rằng trong suốt cuộc đời của một người, mối quan hệ thống kê giữa hành động của một người và đầu vào giác quan là tiêu chí sơ khởi cho việc Học tập kiểu Hebb, hẳn sẽ tạo ra các kết nối synap phù hợp. Đối với trường hợp tái-hướng đến trực tiếp với thính giác hoặc thị giác, điều này không quan trọng, vì âm thanh và tầm nhìn về hành động của chúng ta luôn phù hợp với hành động của chúng ta.
Tuy nhiên, một số hành động có vẻ mờ mịt về mặt nhận thức. Một ví dụ kinh điển là trường hợp nét mặt. Người ta có thể lập luận rằng chúng ta thực sự được sinh ra với các neuron gương để có biểu hiện trên khuôn mặt, dựa trên bằng chứng cho thấy trẻ sơ sinh có nhiều khả năng tạo ra một số biểu cảm khuôn mặt nhất định khi chúng thấy người khác làm như vậy, trước khi việc học có thể tạo ra hiện tượng đó [33]. Mức độ chính xác mà trẻ sơ sinh có thể bắt chước nét mặt vẫn còn là vấn đề tranh luận. Có bằng chứng chắc chắn rằng ít nhất việc thè lưỡi cũng được trẻ sơ sinh bắt chước [34,35], nhưng có ít bằng chứng hơn cho thấy bất kỳ biểu cảm khuôn mặt nào khác đều được bắt chước một cách rõ ràng [31,36,37]. Chúng tôi [6,24,29] và những người khác [31] đã lập luận rằng sự tái-hướng đến gián tiếp có thể cung cấp loại tín hiệu liền kề và ngẫu nhiên cần thiết để huấn luyện các kết nối phù hợp giữa các neuron STS phản ứng với hình ảnh biểu cảm trên khuôn mặt và các chương trình vận động cho thực hiện chúng. Cha mẹ bắt chước nét mặt của trẻ và trẻ trải qua nhiều trường hợp bắt chước trong tương tác trực tiếp với cha mẹ [37]. Do đó, một em bé thường trải qua cảm giác tái-hướng đến gián tiếp khi nhìn/nghe thấy nét mặt của mình bị bắt chước, gây ra những liên hiệp phù hợp với kiểu Hebb. Theo nghĩa này, chúng tôi đề xuất rằng cấu trúc di truyền của chúng ta có thể tạo điều kiện thuận lợi cho sự phát triển của các neuron gương cho các biểu hiện trên khuôn mặt, nhưng không phải (hoặc ít nhất là không nói chung) bằng cách nối trước các kết nối STS-PM để trẻ sơ sinh được trang bị các neuron gương trưởng thành cho nét mặt, mà bằng cách trang bị cho trẻ có xu hướng nhìn chằm chằm vào khuôn mặt của cha mẹ và cha mẹ có xu hướng bắt chước nét mặt của trẻ [29]. Cuối cùng, sự phát triển của các neuron gương cho các biểu hiện trên khuôn mặt khi đó vẫn phụ thuộc vào việc học hỏi trong suốt cuộc đời, nhưng điều này sẽ được điều chỉnh bởi các khuynh hướng hành vi như vậy. Tác động của việc bắt chước đối với việc Học tập kiểu Hebb có lẽ sẽ kém nhanh hơn so với việc tái-hướng đến trực tiếp, vì việc bắt chước nét mặt của chúng ta sẽ thay đổi nhiều hơn về thời gian và đặc tính hình ảnh. Trong thế giới hiện đại của chúng ta, những chiếc gương vật lý cũng có thể góp phần tạo ra sự tái-hướng đến trong trường hợp biểu hiện trên khuôn mặt, và sự tái-hướng đến này sẽ đặc biệt phù hợp với việc Học tập kiểu Hebb, nhưng vẫn chưa rõ vai trò của những chiếc gương vật lý này trong đời sống phát triển điển hình là tới mức nào.
Tuy nhiên, sự phát triển không cần phải luôn là nhìn thấy. Bởi vì trẻ sơ sinh nghe thấy chính mình khóc và cười, nên các neuron phản chiếu thính giác đối với những âm thanh cảm xúc này có thể xuất hiện mạnh mẽ ngay cả khi không có sự bắt chước bằng hình ảnh của cha mẹ. Trong khi bập bẹ, em bé cũng tạo ra các yếu tố ngẫu nhiên trong việc kích hoạt các neuron tiền vận động, kích hoạt giọng nói tạo dựng và các neuron ở thùy thái dương phản ứng với lời nói đó. Khi các kết nối synap đã được huấn luyện bằng tiếng bập bẹ của chính nó, việc nghe cha mẹ nói có thể kích hoạt các chương trình vận động để tái tạo các từ [6]. Quá trình này sẽ được hỗ trợ bởi thực tế là cha mẹ thay đổi giọng nói của chính họ để giống với giọng nói của em bé hơn (người mẹ [38]). Ở đây, xu hướng đa văn hóa của cha mẹ đối với người mẹ và xu hướng bập bẹ của trẻ sơ sinh sẽ tạo điều kiện cho sự xuất hiện của các neuron gương khớp nối thích hợp.
Ngược lại, nhiều kích thích khác không phù hợp với chương trình vận động của chúng ta, thỉnh thoảng xảy ra trong khi chúng ta thực hiện một hành động (hãy tưởng tượng một em bé đang cầm nắm ở một nhà giữ trẻ, với những em bé khác đang bò xung quanh và ném đồ đạc), nhưng những đầu vào cảm giác này sẽ không có cùng khả năng xảy ra hoặc ưu tiên chặt chẽ về mặt thời gian đối với việc kích hoạt các chương trình vận động cụ thể và do đó sẽ giảm trung bình như gây nhiễu như vậy. Tuy nhiên, một số trường hợp đặc biệt có thể tạo ra sự ưu tiên chặt chẽ về mặt thời gian và các tình huống ngẫu nhiên không trùng khớp. Ví dụ, mỗi khi một người đưa thứ gì đó cho em bé, hình ảnh bàn tay đặt đồ vật sẽ có ngay trước khi thực hiện hành động nắm và có thể dẫn đến một mức độ liên hiệp nào đó giữa các neuron STS về việc đặt và neuron PM để nắm bắt. Thật vậy, cái gọi là ‘neuron gương có liên quan về mặt logic’ dường như tồn tại [2], và các thí nghiệm trong phòng thí nghiệm cho thấy rằng việc liên tục gặp phải các tình huống bất ngờ không khớp có thể tạm thời liên hiệp các chương trình vận động với các quan sát hành động không khớp (xem phần 4 của [39] cho tổng quan). Ngoài ra, một vật thể có thể cầm nắm theo một cách cụ thể sẽ luôn hiện diện một cách có hệ thống khi bé cầm vật thể đó theo chính cách đó, dự đoán mối liên hệ kiểu Hebb giữa các neuron hình dạng trong hệ thống thị giác và các neuron PM mã hóa khả năng thực hiện chức năng với vật thể này. Thật vậy, những kết nối như vậy dường như tồn tại và có thể được quan sát theo cái gọi là các neuron kinh điển.
Thật không may, có rất ít công trình kiểm tra bằng thực nghiệm giả định của chúng ta rằng các mối quan hệ thống kê (ngẫu nhiên và xảy ra trong khoảng thời gian tạm thời của việc Học tập kiểu Hebb) giữa những gì chúng ta làm và những gì chúng ta cảm nhận (nghe và nhìn thấy) nói chung là phù hợp với các mối quan hệ trong hệ thống neuron gương sẽ chiếm ưu thế. Một số ít nghiên cứu đã phân tích đoạn phim về trẻ sơ sinh và cha mẹ chúng và phát hiện ra rằng cha mẹ thường bắt chước giọng nói và nét mặt của trẻ, và trẻ sơ sinh được biết là dành nhiều thời gian để nhìn vào bàn tay chính mình và nét mặt của (là thường bắt chước) người chăm sóc chúng (xem [32] và phần 5 của [37] để tổng quan). Một cách hiệu quả để kiểm tra giả thuyết của chúng tôi là ghi lại thông tin đầu vào cảm giác của em bé và hành động của chính em bé trong một khoảng thời gian đáng kể để kiểm tra mối quan hệ thống kê giữa đầu ra của hoạt động và đầu vào thính giác-thị giác. Hiện tại, kiểm soát một dự án như vậy khó có thể thực hiện được là do cần có lao động thủ công để phân tích số ngày ghi chép như vậy. Tuy nhiên, với việc các thiết bị gắn trên đầu (ví dụ: kính Google) và theo dõi chuyển động ba chiều (ví dụ: Kinect của Microsoft) đang trở nên phổ biến, chúng ta có thể sớm định lượng một cách có hệ thống các khả năng cảm biến-vận động mà trẻ sơ sinh thực sự gặp phải. Cho đến lúc đó, phần còn lại của lập luận của chúng tôi chỉ dựa trên giả định rằng các tình huống cảm biến vận động sẽ ủng hộ một tỷ lệ đáng kể các kết nối phù hợp kiểu Hebb giữa STS và PM.
(c) Từ tái-hướng đến đến thuộc tính gương
Nếu các kết nối phù hợp như vậy đã được huấn luyện và cá nhân nghe thấy ai đó thực hiện một hành động tương tự, thì âm thanh của hành động đó, giống với các âm thanh tái-hướng đến có liên quan đến hành động trong quá khứ của người nghe, sẽ kích hoạt các neuron STS, sẽ kích hoạt, thông qua các synap được tăng cường, neuron PM kích hoạt việc thực hiện các hành động tạo ra âm thanh tương tự. Các neuron PM sẽ trở thành neuron gương. Hoạt động của neuron PM trong khi lắng nghe hành động của người khác về cơ bản sẽ là tái hiện về những trí nhớ quy trình trong quá khứ về trạng thái vận động xảy ra cùng với các sự kiện cảm giác này, nhưng là tái hiện được kích hoạt thông qua kích thích xã hội bên ngoài. Điều này đặt các neuron gương vào một nhóm hiện tượng tái kích hoạt rộng hơn, bao gồm cả trí nhớ và trí tưởng tượng.
Trường hợp về trường nhìn phức tạp hơn: hành động của chính một người được nhìn từ góc độ lấy cái tôi làm trung tâm, hành động của những người khác được nhìn từ góc độ khác, lấy sự phân bổ làm trung tâm. Vậy làm thế nào việc nhìn thấy hành động của người khác sẽ kích hoạt các neuron STS phản ứng với việc nhìn thấy hành động của chính chúng ta? Đầu tiên, một số neuron STS phản ứng với hình ảnh của một hành động được nhìn từ một số góc độ khác nhau [24]. Làm thế nào những neuron này có được đặc tính như vậy vẫn chưa hoàn toàn rõ ràng, nhưng ở loài khỉ, quan điểm bất biến như vậy có thể xuất hiện sau khi trải nghiệm những góc nhìn khác nhau của cùng một vật thể ba chiều [41]. Theo đó, đây có thể là cơ hội để nhìn thấy hành động của người khác từ một số góc độ giúp cung cấp cho các neuron STS khả năng phản ứng với việc nhìn thấy hành động qua các góc độ, và do đó là các tác nhân. Thứ hai, neuron có thể biểu thị các đặc tính bất biến về quan điểm nhất định của một hành động (ví dụ: nhịp điệu, tần số thời gian, v.v.), có thể khớp với các hành động có đặc tính tương tự [42] mà không cần phải xoay chuyển hành động trong trường nhìn tâm trí. Thứ ba, các trường hợp bắt chước hoặc gương vật lý sẽ cho phép con người trải nghiệm các loại tình huống ngẫu nhiên có lợi cho việc Học tập kiểu Hebb từ góc nhìn của người thứ ba khi nhìn hành động của người khác nhưng thực hiện hành động của chính họ. Cuối cùng, đối với những hành động có âm thanh đặc trưng, trước tiên các cá nhân có thể trải nghiệm sự ngẫu nhiên giữa việc nhìn và nghe người khác thực hiện những hành động này (ví dụ: nghe thấy lời nói trong khi nhìn thấy chuyển động của môi). Điều này có thể dẫn đến các neuron đa phương thức trong STS [43]. Khi đó, hình ảnh của hành động có thể kích hoạt các hành động vận động phù hợp vì nó kích hoạt hoạt động trong cùng các neuron lời nói nghe-nhìn đã được liên kết với chương trình vận động của người xem trong quá trình tái-hướng đến thính giác. (Sự kết hợp) nào của những hiện tượng này giải thích cho sự xuất hiện của các neuron gương vận động thị giác có thể đối phó với sự khác biệt về quan điểm vẫn còn cần các thí nghiệm nghiên cứu.
(d) Mô tả bổ sung
Dòng chính của các bài báo về hệ thống neuron gương không trực tiếp đề cập đến câu hỏi làm thế nào các neuron gương xuất hiện trong quá trình phát triển nhưng gợi ý rằng những neuron này có thể phục vụ nhận thức xã hội và do đó thúc đẩy khả năng sống sót [2,6,9,25,44–47] . Một số người (ví dụ như Cecilia Heyes) đã đọc những tuyên bố về chức năng như vậy và chỉ ra rằng ‘Quan điểm tiêu chuẩn về MN (neuron gương), mà chúng ta sẽ gọi là “mô tả di truyền”, hợp nhất một tuyên bố về nguồn gốc của MN với một tuyên bố về chức năng của chúng. Nó gợi ý rằng tính phản chiếu của MN chủ yếu là do các yếu tố di truyền có thể di truyền và khuynh hướng di truyền để phát triển MN tiến hóa vì MN tạo điều kiện thuận lợi cho sự hiểu biết hành động [31]. Chúng tôi tin rằng đây là cách đọc không chính xác về công trình khoa học thần kinh về neuron gương: đối với một nhà khoa học thần kinh, việc tuyên bố rằng neuron gương có thể đóng góp vào nhận thức xã hội và do đó mang lại cho động vật những lợi thế về thể chất, không tự động chuyển thành gợi ý rằng con người và khỉ là những mạch nối cứng với nhau để có neuron gương, và việc học đó chỉ có tác động tối thiểu đến neuron gương. Ý của các nhà khoa học thần kinh là nếu một người làm xáo trộn chức năng của các neuron gương, điều này sẽ dẫn đến suy giảm nhận thức xã hội và ngày càng có nhiều bằng chứng ủng hộ tuyên bố này [48]. Tuyên bố như vậy tương thích với bộ gen chịu áp lực chọn lọc để tạo điều kiện thuận lợi cho các neuron gương, nhưng nó không ngụ ý rằng sự lựa chọn này đã tạo ra mã hóa di truyền mạnh hoặc ảnh hưởng di truyền có dạng nối mạch trước khi sinh. Như đã mô tả ở trên, bộ gen có thể cơ sở hóa cho việc học tập của các neuron gương theo kiểu Hebb thay vì tạo khuynh hướng sẵn cho các cá nhân về kiểu đúng của cơ hội học tập [29]. Nói tóm lại, ‘quan điểm tiêu chuẩn’ có thể bị chỉ trích vì đã bỏ qua quá trình hình thành bản thể của các neuron gương nhưng không cho rằng các neuron gương với tất cả sự phức tạp của chúng đều được mã hóa về mặt di truyền và miễn nhiễm với việc học.
Trong số các lý thuyết đề cập đến sự hình thành bản thể của neuron gương, tất cả dường như đều cho rằng trải nghiệm có một vai trò rất quan trọng. Điều này đúng với mô hình Học tập kiểu trường phái Hebb của chúng tôi, học theo trình tự liên hiệp [28,30,31], mô hình của Casile et al. [34], mô hình biểu sinh của Ferrari và cộng sự [49], mô hình Bayes của Kilner và cộng sự [50] và cho phần lớn các mô hình tính toán của hệ thống neuron gương [51]. Tất cả các mô hình cũng đóng vai trò quan trọng đối với khuynh hướng di truyền, ít nhất là trong việc kết nối các vùng cảm giác và vận động với các kết nối synap và triển khai một số quy tắc học tập cơ bản vào hệ thống. Sự khác biệt chính giữa các mô hình có lẽ là mức độ mô tả mà chúng hướng tới trực tiếp nhất. Mô hình học tập kiểu Hebb của chúng tôi là một phương pháp tiếp cận khoa học thần kinh từ dưới lên, bắt đầu từ các khối xây dựng nhỏ của hệ thống—độ khả biến phụ thuộc-thời gian-nhọn sóng xảy ra ở các synap và các chi tiết giải phẫu của các kết nối (xem thêm mục sau)—và kiểm tra xem ở các neuron gương có xuất hiện từ dưới lên sự tương tác của các khối xây dựng này hay không. Học theo trình tự liên hiệp không phải là mô hình thần kinh mà là mô hình nhận thức và nhấn mạnh các biến cấp hệ thống mà các thí nghiệm hành vi đã cho thấy là rất quan trọng đối với việc học liên hiệp, nhưng không đề cập đến cách thức thực hiện việc học trong sinh học của các synap [28,31]. Casile và cộng sự, [34] cảnh báo chúng ta về khả năng các neuron gương cho các hành động khác nhau có thể xuất hiện theo những cách khác nhau: sự chuẩn bị trước về mặt di truyền có thể quan trọng hơn đối với nét mặt, trong khi việc Học tập kiểu Hebb có thể quan trọng hơn đối với các hành động bằng tay [34]. Mô hình biểu sinh cho biết thêm rằng kinh nghiệm có thể tác động không chỉ bằng cách kích hoạt quá trình Học tập kiểu Hebb mà còn bằng cách sửa đổi về mặt biểu sinh phần nào của gen có thể được biểu hiện [49]. Các mô hình tính toán nhấn mạnh kiến trúc tổng thể của hệ thống về mặt nội dung thông tin nhưng thường sử dụng thuật toán lan truyền ngược lỗi mà không có giả thuyết cụ thể nào về việc triển khai sinh học các quy tắc học này [51]. Cuối cùng, mô tả mã hóa dự đoán của Kilner và cộng sự [50] mô tả các neuron gương ở cấp độ hệ thống bằng cách sử dụng thống kê kiểu Bayes. Làm thế nào những số liệu thống kê này được tính toán trong sinh học của các synap không nằm trong phạm vi lý thuyết của Kilner và cộng sự.
Do đó, chúng tôi cảm thấy rằng có sự đồng thuận cơ bản về tầm quan trọng của việc học trong quá trình hình thành neuron gương. Việc mô tả lĩnh vực này được tạo thành từ hai phe, trong đó một phe được cho là cho rằng tất cả đều là do di truyền và phe kia tạo ra các thí nghiệm để bác bỏ giả thuyết di truyền, có vẻ như là một sự bóp méo. Thay vào đó, các lý thuyết hiện tại dường như không có nhiều lựa chọn thay thế mang tính cạnh tranh, mà là những nỗ lực song song nhằm khám phá cách trải nghiệm có thể tạo ra một hiện tượng rất phức tạp bắt đầu từ các cấp độ tập trung khác nhau, với một số tập trung vào thấp nhất, cấp độ synap, một số khác tập trung vào sự tương tác giữa các vùng não và số khác vẫn ở cấp độ liên hiệp của các thực thể nhận thức. Trong những thập kỷ tiếp theo, thách thức lớn nhất sẽ là việc thống nhất những nỗ lực mang tính “khu trú” này thành một mô hình thống nhất dành cho tất cả các cấp độ. Cùng lúc, việc tranh luận xem cách tiếp cận nào là tốt nhất dường như chẳng có tác dụng gì. Việc để các mô hình khác nhau phát triển hơn nữa sẽ làm sáng tỏ các cấp độ mà mỗi mô hình khám phá một cách trực tiếp nhất. Theo tinh thần này, giờ đây chúng tôi chuyển sang khám phá một chiều thời gian tốt hơn của việc Học tập kiểu Hebb để cho thấy các dự đoán từ dưới lên synap ăn khớp như thế nào với các dự đoán từ trên xuống hơn được thực hiện bởi Kilner et al. [50].
GÓC NHÌN VÀ DỰ ĐOÁN KIỂU HEBB THEO THỜI GIAN-VI MÔ
Một đặc điểm chính trong sự hiểu biết hiện đại của chúng ta về việc Học tập kiểu Hebb là sự nhạy cảm tinh tế của nó đối với các mối quan hệ tạm thời khá tốt đẹp của hoạt động trước và sau synap. Do đó ở đây chúng tôi kiểm tra ý tưởng cốt lõi về mô tả học tập kiểu Hebb của chúng tôi—sự liên hệ lại trực tiếp hoặc gián tiếp—ở khoảng thời gian mili giây này, mở rộng mô tả ngắn gọn được trình bày trong Keysers và Keysers và cộng sự [6,52].
(a) Dự đoán chuyển tiếp kết nối
Nếu bạn nghĩ đến việc với lấy một chiếc bánh quy, nắm lấy nó rồi đưa vào miệng, thì ở thế giới bên ngoài, thời điểm của từng thành phần phụ của hành động và hậu quả cảm giác của chúng trùng khớp chính xác về mặt thời gian (hình 3a). Tuy nhiên, phải mất khoảng 100 ms để hoạt động tiền vận động kích hoạt các hành động công khai phức tạp như với tới và nắm lấy [53]. Sau đó, phải mất thêm 100 ms để âm thanh/hình ảnh của hành động đó kích hoạt hoạt động trong STS [54]. Do đó, điều này sẽ làm thay đổi tốc độ của các neuron STS đại diện cho hình ảnh và âm thanh của một hành động khoảng 200 ms so với tốc độ của các neuron PM đã kích hoạt hành động đó (hình 3b). Do đó, khái niệm thời gian-vĩ mô cho rằng hoạt động trong các neuron STS cho một hành động trùng lặp về thời gian với hoạt động của các neuron PM kích hoạt hành động đó thực sự là một sự đơn giản hóa quá mức. Điều này gây ra hậu quả đối với việc Học tập kiểu Hebb, bởi vì phản ứng STS đối với ngữ cảnh đạt được không còn xảy ra ngay trước hoạt động tiếp cận của các neuron PM, như yêu cầu của cửa sổ 40 ms về tính khả biến phụ thuộc-thời gian-nhọn sóng (hình 1). Thay vào đó, việc kích hoạt các neuron trong STS phản ứng với một giai đoạn cụ thể của hành động (ví dụ: tiếp cận) trước hoạt động thần kinh PM kích hoạt giai đoạn tiếp theo (ví dụ: nắm bắt) và việc Học tập kiểu Hebb chủ yếu sẽ củng cố các kết nối giữa STS tiếp cận và các neuron nắm bắt PM. Do đó, kết quả học tập nổi trội phải là mối liên hệ với các đặc tính dự đoán. Một số hoạt động Học tập kiểu Hebb vẫn có thể xảy ra trong một giai đoạn hành động nhất định, bởi vì các đợt tăng đột biến sớm của STS tiếp cận các neuron thần kinh xảy ra ngay trước khi các đợt tăng đột biến muộn của PM đến các neuron.
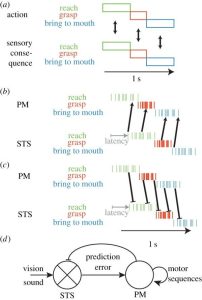
Hình 3. (a) Trong thế giới thực, việc thực hiện một hành động cũng như hình ảnh và âm thanh của từng giai đoạn của một hành động xảy ra cùng một lúc và do đó người ta có thể dự đoán rằng các giai đoạn tương ứng trong miền cảm giác và vận động sẽ được liên kết. (b) Thay vào đó, độ trễ làm thay đổi các phản ứng trong STS so với các neuron tiền vận động (PM) và việc Học tập kiểu Hebb ở quy mô thời gian tốt dự đoán mối liên hệ giữa các giai đoạn tiếp theo, tức là các dự đoán. (c) Phản hồi ức chế từ PM đến STS cũng phải tuân theo việc Học tập kiểu Hebb và tạo ra các lỗi dự đoán trong hệ thống động mới nổi (d). (Phiên bản trực tuyến có màu.)
Hệ thống này sẽ dự đoán được bao nhiêu? Nếu chúng ta có độ trễ tạm thời khoảng 200 ms giữa hoạt động của neuron PM và việc kích hoạt các neuron STS đại diện cho sự tái quan hệ, thì việc nhìn thấy một thành phần hành động xảy ra ở thế giới bên ngoài tại thời điểm t sẽ kích hoạt hoạt động (thông qua các synap được được định hình bằng cách học kiểu Hebb) trong neuron PM đại diện cho thành phần hành động thường xảy ra ở thế giới bên ngoài với tốc độ t + 200 ms. Do đó, độ trễ vận động và cảm giác xác định trực tiếp phạm vi dự đoán của kết nối cảm biến-vận động. Vậy thì, việc Học tập kiểu Hebb sẽ đào tạo một hệ thống dự đoán chỉ đơn giản là do sự bất đối xứng về thời gian của STDP (hình 1) và độ trễ đã biết trong hệ thống cảm giác và vận động (hình 3b).
Trong thế giới thực, các thành phần hành động có thể tổ chức thành nhiều chuỗi hành động khác nhau như các chữ cái trong từ và các kết nối STS → PM mang tính dự đoán này có thể phản ánh sự phân bố xác suất chuyển đổi của các hành động của chúng ta: nếu trong lịch sử vận động trước đây của chúng ta, hành động A chưa bao giờ theo sau là hành động x1 (p(x1|A) = 0), đôi khi là x2 (p(x2|A) = 0.2) và thường là x3 (p(x3|A) = 0.8), việc Học tập kiểu Hebb sẽ mong đợi một STS neuron phản ứng với A để có trọng số kết nối gần như bằng 0 với các neuron PM kích hoạt ×1, trọng số 0,2 với các neuron kích hoạt ×2 và trọng số 0,8 với các neuron kích hoạt ×3. Do đó, neuron PM cho ba hành động này phải có trạng thái hoạt động là 0, 0.2 và 0.8 theo biểu hiện của hành động A trong STS. Khi đó, mô hình hoạt động trong PM là phân bố xác suất của các hành động tiếp theo phản ánh các khả năng vận động ngẫu nhiên trong quá khứ của người quan sát và có thể đóng vai trò là yếu tố đi trước (theo nghĩa Bayes) cho hành động có khả năng xảy ra tiếp theo.
(b) Kết nối ngược bị ức chế và lỗi dự đoán
Một yếu tố thường bị bỏ qua trong giải phẫu của hệ thống neuron gương là sự hiện diện của các kết nối ngược từ PM đến STS, dường như có ảnh hưởng ức chế thuần [55,56]. Theo quan điểm của Hebb, đối với những kết nối này, tình huống hơi khác một chút, vì các neuron PM thực sự hoạt động trước các neuron STS, như việc Học tập kiểu Hebb yêu cầu, mặc dù 200 mili giây thay vì 40 mili giây trước đó là tối ưu cho việc Học tập kiểu Hebb. Do đó, đối với các kết nối phản hồi ức chế này, các dự đoán ức chế từ neuron PM mã hóa một giai đoạn cụ thể của hành động phải được tăng cường bằng các biểu diễn STS trên cùng một hành động và hành động xảy ra ngay trước đó (hình 3c).
Khi chúng ta xem xét cả luồng thông tin tiến và lùi, hệ thống neuron gương dường như không còn là một hệ thống liên kết đơn giản trong đó việc nhìn thấy một hành động nhất định sẽ kích hoạt sự thể hiện vận động của hành động đó. Thay vào đó, nó trở thành một hệ thống động (hình 3d). Hình ảnh và âm thanh của một hành động sẽ kích hoạt hoạt động trong các neuron STS. Điều này dẫn đến một mô hình kích hoạt dự đoán của các neuron PM mã hóa hành động xảy ra sau 200 ms so với những gì neuron STS đại diện, với mức độ kích hoạt tương ứng của chúng biểu thị khả năng xảy ra của chúng dựa trên các tình huống cảm biến vận động trong quá khứ. Tuy nhiên, hệ thống sẽ không dừng lại ở thời điểm đó. Dự đoán này trong neuron PM được gửi ngược lại dưới dạng tín hiệu ức chế đến neuron STS. Bởi vì phản hồi phải được gửi đến các neuron đại diện cho các hành động trước đó và hiện tại được thể hiện trong PM, nên nó sẽ gây ra hai hậu quả. Nó sẽ chấm dứt sự thể hiện cảm giác của các hành động trong quá khứ, điều này có thể góp phần vào cái thường được gọi là đoạn che lấp ngược/ backward masking trong tư liệu thị giác [57]. Thứ hai, bằng cách hủy bỏ các biểu hiện liên quan đến x1, x2 và x3 với xác suất tương ứng của chúng, về cơ bản nó sẽ ức chế các neuron STS đại diện cho các hậu quả cảm giác dự kiến của hành động mà neuron PM dự đoán sẽ xảy ra. Ở cấp độ khái niệm hơn, nó sẽ ức chế giả thuyết rằng các neuron PM thích thú về hành động tiếp theo sẽ được nhận thức. Khi bộ não nhìn và nghe thấy hành động nào thực sự xảy ra tiếp theo, nếu đầu vào này phù hợp với giả thuyết, thì hậu quả cảm giác của hành động đó sẽ bị ức chế một cách tối ưu và sẽ có ít thông tin được gửi từ STS → PM. Bởi vì các neuron PM (và các neuron đỉnh sau [58]) được tổ chức thành chuỗi hành động trong vỏ não tiền vận động, nên việc thể hiện hành động x3 sau đó sẽ kích hoạt việc kích hoạt những hành động thường theo sau hành động x3 trong quá trình thực hiện, hoạt hóa tạo ra toàn bộ chuỗi hành động sự biểu diễn của các neuron PM mà không cần thêm bất kỳ thúc đẩy cảm giác nào và những dự đoán tiếp theo này sẽ tiếp tục ức chế đầu vào STS trong tương lai. Nếu hành động x2 diễn ra sau hành động A, sự ức chế sẽ yếu hơn và nhiều biểu hiện cảm giác của x2 sẽ rò rỉ qua PM. Điều này sẽ thể hiện một ‘lỗi dự đoán’, lỗi này sẽ thay đổi mô hình hoạt động PM để phù hợp hơn với đầu vào, khác với những kỳ vọng trước đó. Nếu hành động x1 diễn ra sau hành động A thì sẽ không có sự hủy bỏ nào được áp dụng trong STS và hoạt động mạnh nhất sẽ được gửi từ STS → PM, định tuyến lại hoạt động PM vào luồng hành động thường theo sau x1, thay vì x3, như ban đầu giả thuyết.
Ở độ phân tích tạm thời này, trong quá trình quan sát/nghe hành động, mô hình hoạt động giữa các nút trong PM không còn là sự phản ánh đơn giản về những gì xảy ra trong STS mà là sự phân bổ xác suất được dự đoán tích cực về những gì người quan sát sẽ cảm nhận được cá nhân được quan sát sẽ làm gì tiếp theo. Nhờ Học tập kiểu Hebb, toàn bộ vòng lặp STS-PM trở thành một hệ thống động thực hiện mã hóa dự đoán. Khi hành động được quan sát diễn ra hoàn toàn như mong đợi, hoạt động trong PM sẽ thực sự được tạo ra bằng cách sử dụng trình tự của điều khiển vận động thông thường thay vì bằng đầu vào thị giác.
(c) Quy luật học tập và điểm liên hệ với các mô hình khác
Một hệ quả quan trọng của sự ức chế phản hồi cũng là trong quá trình tái-hướng đến, một khi hệ thống đã học được, việc thực hiện một hành động sẽ kích hoạt phản hồi ức chế đối với các neuron STS để đảm bảo rằng các neuron STS thực sự hạn chế đầu vào của chúng đối với các neuron PM gây ra hành động, và việc Học tập kiểu Hebb sẽ tự giới hạn. Nếu các tình huống ngẫu nhiên thay đổi, ví dụ: một người học một kỹ năng mới như chơi piano, neuron PM không dự đoán được sự tái-hướng đến của thính giác và quá trình học tập mới xảy ra do đầu vào mới từ STS được gửi đến neuron PM có tiềm năng/ điện thế Học tập kiểu Hebb.
Việc tính toán các lỗi dự đoán trong hệ thống học tập kiểu Hebb này tạo ra một điểm kết nối quan trọng với các mô hình khác của hệ thống neuron gương. Mô hình mã hóa dự đoán của Kilner và cộng sự [50] không chỉ ra cách não thực hiện các dự đoán Bayes trong các synap của nó, nhưng đề xuất rằng hoạt động PM thể hiện ước tính Bayes về các hành động trong tương lai, cho phép người quan sát suy ra ý định vận động của cá nhân được quan sát. Mô hình này coi luồng thông tin STS → PM chỉ là tín hiệu cập nhật cho xác suất Bayesian của các trạng thái tiền vận động. Mô hình của chúng tôi đưa ra những cách giải thích rất giống nhau từ góc độ từ dưới lên. Do đó, mô hình Học tập kiểu Hebb của chúng tôi có thể bổ sung cho mô hình dự đoán Bayes bằng cách triển khai sinh học từ dưới lên hợp lý. Đổi lại, mô hình dự đoán Bayes giúp diễn giải quá trình xử lý thông tin mà chúng ta mô tả dưới góc độ của cái có thể được gọi là cuộc cách mạng mã hóa dự đoán hoặc Bayes trong khoa học não bộ. Thật vậy, nhiều miền khoa học não bộ hiện nay đã ngừng coi tri giác là một quá trình có thứ bậc trong đó thông tin cảm giác được gửi đi một cách thụ động từ các vùng não thấp hơn đến cao hơn. Thay vào đó, tri giác ngày càng được coi là một quá trình có tác động tích cực hơn, trong đó não đưa ra các dự đoán dựa trên kinh nghiệm trong quá khứ (tương đương với các xác suất trước đó theo thuật ngữ Bayes), được gửi từ các vùng não cao hơn đến thấp hơn trong hệ thống phân cấp và được trừ ra khỏi đầu vào giác quan thực tế. Đầu vào cảm giác được gửi từ vùng thấp hơn đến vùng cao hơn sau khi trừ đi các dự đoán, khi đó sẽ là một lỗi dự đoán dùng để cập nhật các dự đoán, thay vì trực tiếp thúc đẩy tri giác. Khuôn khổ rất chung này đã được sử dụng rất thành công để hiểu hoạt động thần kinh ở giai đoạn đầu của vỏ não thị giác [59,60] nhưng gần đây cũng đã được sử dụng để khái niệm hóa hệ thống neuron gương [50] và thậm chí cả tâm trí hóa [61].
Bằng chứng về việc mã hóa dự đoán trong hệ thống neuron gương vẫn còn hiếm nhưng đã bắt đầu xuất hiện. Bản chất dự đoán của phản ứng PM được thể hiện rõ ràng từ thực tế là hình ảnh đạt được làm tăng tính dễ bị kích thích của các cơ liên quan đến giai đoạn hành động có khả năng xảy ra nhất là nắm bắt [62]. Khả năng hoạt động PM có thể được điều khiển bởi các dự đoán bên trong trong trường hợp không có đầu vào thị giác rõ ràng xuất phát từ quan sát rằng các neuron gương phản ứng trong quá trình thực hiện hành vi nắm bắt phản ứng với cảnh chạm vào phía sau màn hình mờ [20] và neuron gương thính giác đó phản ứng với âm thanh nứt của đậu phộng được bóc vỏ bắt đầu truyền dẫn trước cả khi xem các bàn tay đang nắm lấy hạt đậu phộng [3]. Bằng chứng cho thấy các dự đoán từ PM → STS hủy bỏ các hành động được dự đoán và do đó làm im lặng luồng thông tin STS → PM khi và chỉ khi các hành động đó có thể dự đoán được, xuất phát từ thực tế là hướng chủ yếu của luồng thông tin là từ PM → STS khi quan sát các hành động có thể dự đoán được, nhưng STS → PM khi quan sát sự khởi đầu không thể đoán trước của một hành động [63].
(d) Học tập kiểu Hebb và tham gia hành động chung
Con người có thể cùng nhau hành động với độ chính xác đáng kinh ngạc về mặt thời gian. Các nghệ sĩ piano trong một bản hợp tấu có thể đồng bộ hóa hành động của họ trong vòng 30 mili giây kể từ người nhạc trưởng đưa ra dấu hiệu [64]. Với độ trễ cảm biến vận động khoảng 200 ms mà chúng tôi đã đề cập ở trên, làm sao điều này có thể xảy ra? Chẳng phải phải mất 200 mili giây để một nhạc sĩ nghe được người chỉ huy đưa dấu hiệu chơi và phản hồi lại điều đó sao? Một trong những ý nghĩa mạnh mẽ của việc phân tích chi tiết về việc Học tập kiểu Hebb là vì các kết nối synap được huấn luyện bằng sự tái-hướng đến bao gồm độ trễ cảm giác vận động điển hình của con người, nên chúng huấn luyện các kết nối từ STS → PM để thực hiện các dự đoán về tương lai trong một khoảng dịch chuyển thời gian/ time-shift sẽ bù đắp độ trễ cảm biến vận động gặp phải khi hành động với một cá nhân khác có độ trễ tương tự. Điều này là do sẽ mất khoảng thời gian tương tự (khoảng 200 ms) để chương trình vận động của bạn kích hoạt các neuron STS, cũng như để chương trình vận động của bạn kích hoạt các neuron STS của tôi khi tôi đang nghe/quan sát bạn. Do đó, việc Học tập kiểu Hebb, bằng cách tái liên kết đào tạo các dự đoán cảm biến vận động cho phép các hành động chung chính xác mặc dù độ trễ cảm biến vận động kéo dài.
(e) Học tập kiểu Hebb và truyền dẫn
Một hệ quả quan trọng của quan điểm cho rằng hệ thống neuron gương được kết nối dựa trên tái-hướng đến là não liên hiệp các trạng thái bên trong hiện diện khi chúng ta thực hiện một hành động nhất định với âm thanh và hình ảnh của hành động đó. Theo đó, khi chúng ta chứng kiến hành động của người khác, kiểu hoạt động vận động sẽ được kích hoạt theo dự đoán ở nhân chứng không hẳn là sự phản ánh những gì xảy ra trong não của người thực hiện mà là sự truyền dẫn những gì đã xảy ra trong não của chính chúng ta, khi chúng ta thực hiện những hành động như vậy. Bởi vì con người chia sẻ khoảng 99% gen của họ với những người khác và có lẽ hơn 90% gen với khỉ, giả thiết rằng các trạng thái vận động ẩn xảy ra trong hành động của chính chúng ta là một mô hình phù hợp cho những trạng thái xảy ra trong não của người hoặc khỉ khác thì không phải là vô lý. Nó cấu thành một thông tin ‘trước’ có thể được cập nhật bằng các bằng chứng trái ngược nếu có. Tuy nhiên, người quan sát càng khác biệt với tác nhân được quan sát thì bản chất phóng chiếu của quá trình này càng trở nên rõ ràng.
Để kiểm tra dự đoán này, chúng tôi đã đo hoạt động của não bằng cách sử dụng hình ảnh cộng hưởng từ chức năng (fMRI) trong ba điều kiện [65]. Những người tham gia thực hiện các hành động bằng tay (ví dụ: xoay ly rượu). Họ nhìn thấy một người khác thực hiện những hành động tương tự. Cuối cùng, họ nhìn thấy một robot công nghiệp thực hiện những hành động tương tự. Việc nhìn thấy con người thực hiện hành động (hình 4a) đã kích hoạt một mạng lưới hoạt động của não bộ, vùng tiền vận động và não đỉnh (hình 4b) tương tự như mạng lưới được những người tham gia sử dụng để thực hiện hành động tương tự (hình 4c). So sánh mô hình hoạt động của người quan sát và người thực thi (b–c) cho thấy sự tương đồng đáng kể (r(b,c) = 0,5)—bộ não đã thành công trong việc mô phỏng chính xác hoạt động não của tác nhân. Tuy nhiên, khi người tham gia xem robot thực hiện hành động (hình 4d), họ đã tạo ra một mô hình hoạt động của não (hình 4e) không giống như hoạt động của bộ xử lý khiến robot di chuyển (hình 4f). Thay vào đó, mô hình tiếp tục giống với thứ mà người tham gia sẽ sử dụng để thực hiện hành động này (hình 4c). Điều này minh họa bản chất phóng chiếu của việc đọc suy nghĩ thông qua hệ thống neuron gương.
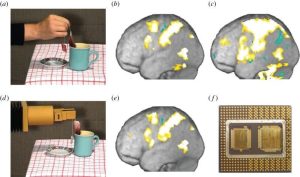
Hinh 4. Việc nhìn thấy con người thực hiện một hành động (a) dẫn đến các kích hoạt trong hệ thống gương (b) giống với hoạt động trong quá trình con người thực hiện các hành động tương tự (c). Việc nhìn thấy robot thực hiện các hành động tương tự (d) sẽ tạo ra mô hình hoạt động trong hệ thống gương (e) rất khác với mô hình hoạt động khiến robot hành động (f), nhưng giống với mô hình mà người xem sẽ sử dụng thực hiện một hành động tương tự (c). Các bảng (a-e) được điều chỉnh từ Gazzola và cộng sự [65].
BÊN NGOÀI HỆ THỐNG VẬN ĐỘNG
Bởi vì các neuron gương lần đầu tiên được tìm thấy ở PM [1–4,20] và ở các vùng đỉnh sau [21,58], nơi kiểm soát các hành động, các khía cạnh vận động của việc đọc suy nghĩ được chú ý nhiều hơn. Nhưng bằng chứng từ một số nguồn hiện nay cho thấy rằng phần cao nhất của vỏ não cảm giác-cơ thể sơ cấp cũng được kích hoạt gián tiếp khi chúng ta nhìn thấy hành động của người khác và vỏ não cảm giác-cơ thể thứ cấp hoạt động khi chúng ta thấy người khác bị chạm vào [45]. Ngoài ra, các vùng liên quan đến trải nghiệm cảm xúc cũng được kích hoạt gián tiếp khi chúng ta chứng kiến những người khác trải qua những cảm xúc tương tự [6,66], bao gồm vùng thùy đảo cho cảm giác ghê tởm, đau và dễ chịu [67–69], vùng mỏm hồi đai cho đau [68] và thể vân cho tưởng thưởng [70].
Chúng tôi vẫn thiếu các bản ghi tế bào-đơn lẻ chứng minh rằng các kích hoạt cảm xúc và cảm giác cơ thể gián tiếp trong fMRI là do các tế bào đơn lẻ phản ứng với cả trải nghiệm và quan sát về cảm giác cơ thể và cảm xúc (nhưng xem [71]). Tuy nhiên, từ góc độ học tập kiểu Hebb, việc có các neuron giống-gương đối với cảm giác cơ thể hoặc cảm xúc không có gì đáng ngạc nhiên. Bất cứ khi nào có thứ gì đó chạm vào da, chúng ta thấy cơ thể mình được chạm vào và chúng ta cảm nhận được sự kích thích giác quan cơ thể. Không giống như trong hệ thống cảm biến vận động, trong đó hoạt động vận động xảy ra trước sự tái-hướng đến thị giác/thính giác, trong trường hợp cảm giác được chạm vào, cả tín hiệu xúc giác và thị giác/thính giác sẽ bị ảnh hưởng bởi độ trễ tương tự so với sự kiện bên ngoài. Do đó, các nhọn sóng từ các neuron thị giác/thính giác và cảm giác cơ thể sẽ tự nhiên rơi vào các cửa sổ thời gian hẹp của quá trình Học tập kiểu Hebb và sẽ củng cố mối liên hệ giữa các neuron mã hóa cảm giác xúc giác bên trong của chúng ta ở S2 với những tế bào mã hóa những gì nhìn và nghe khi sờ chạm ở các vùng giống như STS. Khi nhìn/nghe thấy người khác chạm, những kết nối này có thể kích hoạt hoạt động giống-gương trong S2 và phản chiếu cảm giác chạm của chúng ta lên người mà chúng ta nhìn thấy. Bởi vì dự đoán trong hệ thống STS → PM là do sự khác biệt về độ trễ giữa các neuron này, vốn là nhỏ giữa STS và S2, nên chúng tôi mong đợi các kết nối STS-S2 hiển thị ít mã hóa dự đoán. Tuy nhiên, nếu cảm giác xúc giác là kết quả của các hành động có thể được dự đoán bởi hệ thống STS → PM, thì những dự đoán đó có thể được vi tính toán.
Tương tự như vậy, khi chúng ta chủ động khám phá các vật thể bằng tay, hoạt động trong các neuron PM kiểm soát hành động sẽ diễn ra trước hoạt động không chỉ ở các neuron STS xem và nghe thấy hành động mà còn cả hoạt động của các neuron trong BA2, mã hóa cảm giác xúc giác trải qua khi chạm vào. Do đó, chúng tôi mong đợi sự xuất hiện của một hệ thống động lực giống như trong Hình 1, không chỉ bao gồm STS và PM mà còn cả BA2. Trong hệ thống này, việc Học tập kiểu trường phái Hebb sau đó cũng có thể giải thích cách mọi người học cách ngăn chặn cảm giác xúc giác tự gây ra, tạo ra các lỗi dự đoán xúc giác rất quan trọng đối với việc điều khiển vận động [72], và do đó tại sao bạn không thể tự cù mình [73].
Cuối cùng, đối với cảm xúc, nhiều neuron cũng sẽ được kết nối theo kiểu Hebb. Nếu chúng ta cảm thấy đau đớn vì chị gái vô tình lấy đồ chơi của chị đánh chúng ta, chúng ta thấy đồ chơi đó đánh mình, chúng ta cảm thấy đau, chúng ta nhăn mặt, khóc và bố mẹ chúng ta sẽ phản ánh nét mặt đó. Hình ảnh của cú đánh đi trước nỗi đau của chúng ta, đi trước nét mặt và tiếng khóc mà chúng ta tạo ra, đi trước sự bắt chước khuôn mặt của cha mẹ chúng ta. Nếu lý thuyết của chúng tôi đúng, điều này có thể dẫn đến một chuỗi liên kết kiểu Hebb trên khắp các neuron đại diện cho các trạng thái này. Sau đó, khi chúng ta nhìn thấy hoặc nghe thấy ai đó nhăn mặt vì đau, âm thanh và tầm nhìn sẽ kích hoạt các chương trình vận động phù hợp trên khuôn mặt của chúng ta, từ đó sẽ kích hoạt cảm xúc bên trong của chúng ta [74]. Nếu chúng ta thấy ai đó bị đánh, chúng ta sẽ gián tiếp huy động vỏ não cảm giác và cảm xúc của mình. Và tất cả những hoạt động gián tiếp này sẽ là kết quả của tính khả biến synap trong trải nghiệm của chính chúng ta. Chúng sẽ liên kết những sự kiện có thể quan sát được với những gì chúng ta đã cảm nhận và thực hiện trong những tình huống này. Khi áp dụng chúng cho người khác, chúng ta sẽ phóng chiếu trạng thái của mình, với tất cả những thành kiến vị kỷ không thể tránh khỏi mà điều này sẽ dự đoán.
KẾT LUẬN TỔNG THỂ
Khi hai thập kỷ trước, các neuron gương lần đầu tiên được báo cáo, chúng đã tạo ra một tầm nhìn trong đó hệ thống vận động đóng vai trò đặc biệt trong việc đọc được suy nghĩ của những người khác thông qua nhận thức thể hiện [75]. Ở đây, chúng tôi đề xuất rằng những gì chúng tôi biết về tính khả biến synap phụ thuộc-thời gian-nhọn sóng sẽ định hình sự hiểu biết hiện đại của chúng tôi về việc Học tập kiểu Hebb và cung cấp một khuôn khổ để giải thích, không chỉ cách các neuron gương có thể thể hiện, mà còn cả cách chúng trở nên có các đặc tính dự đoán có thể cho phép đồng bộ gần như-hành động chung. Chúng tôi chứng minh rằng điều này có thể tạo ra một hệ thống có thể cung cấp giải pháp gần đúng cho vấn đề nghịch đảo trong việc suy ra các trạng thái bên trong tiềm ẩn của người khác từ những thay đổi có thể quan sát được trên thế giới thực, nhưng giải pháp này là một phép chiếu bị cản trở bởi những thành kiến ích kỷ. Chúng tôi cũng chỉ ra rằng các neuron gương có lẽ là một trường hợp đặc biệt của các kích hoạt gián tiếp mà dữ liệu fMRI và Học tập kiểu Hebb gợi ý cũng có thể áp dụng cho cách chúng ta chia sẻ cảm xúc và cảm giác với người khác.



- BS Đỗ Thị Thuý Anh dịch và chỉnh lý