
Màng mí mắt (mí mắt thứ ba): Quá trình tạo phản xạ điều kiện (điều kiện hóa) và giải trừ cổ điển ở thỏ Albino



- Bs Đỗ Thị Thúy Anh
Tóm tắt. Sự phân bố độ trễ đáp ứng và đường cong hiệu suất phần trăm của nhóm tạo điều kiện cổ điển, khi so sánh với nhóm đối chứng, chỉ ra rằng sự mở rộng của mí mắt ba (nictitating membrane) được gợi ra bởi một luồng không khí đến giác mạc đã được tạo điều kiện thành công với kích thích trung tính trước đó.
Màng mí mắt (ở người là màng bán nguyệt/plica semilunaris là một nếp hẹp, có nhiều mạch máu, hình lưỡi liềm của kết mạc, nằm ở phía bên và một phần dưới cuống kết mạc; Đường viền bên của nó tự do và tách biệt khỏi kết mạc nhãn cầu, trông giống cùng kết cấu mô học. Biểu mô của plica rất giàu tế bào cốc; Lớp đệm của plica chứa chất béo và một số cơ không có vân) ở thỏ bạch tạng bao gồm một tấm sụn cong được bao phủ bởi biểu mô tuyến. Nó được kéo từ khóe mắt bên trong của mắt ngang qua giác mạc bằng một tấm cơ trơn, nhưng cơ chế hoạt động của nó vẫn chưa được hiểu rõ ràng (7).
Mặc dù chưa có nỗ lực nào được báo cáo nhằm tạo điều kiện màng này ở thỏ, nhưng những quan sát ngẫu nhiên trong phòng thí nghiệm của chúng tôi đã chỉ ra rằng sự mở rộng của màng được tạo ra một cách đáng tin cậy bằng một luồng không khí thổi vào giác mạc. Chúng tôi đã quan sát thấy rằng khi màng được kích hoạt, nó hiếm khi kéo dài quá đường giữa của đồng tử và luôn để lộ một phần giác mạc, và ngay cả những con thỏ được tạo phản xạ điều kiện tốt cũng dường như không có khả năng mở rộng màng một cách bền vững. Hai đặc tính này dường như mang lại cho người thực nghiệm mức độ kiểm soát thậm chí còn cao hơn đối với các hậu quả cảm giác của kích thích không điều kiện so với đặc tính hiện có trong nghiên cứu được báo cáo trước đây của chúng tôi về tạo phản xạ điều kiện mí ngoài của thỏ (2).
Thiết bị tạo phản xạ điều kiện và cách thức nhốt thỏ trong hộp Plexiglas đã được mô tả (2, 3). Để cho phép ghi lại chuyển động của màng và đảm bảo giác mạc tiếp xúc liên tục, mí mắt trên và dưới của mắt phải của thỏ được dán mở. Khi hộp hạn chế được đặt trong một thiết bị làm lạnh không hoạt động, một chiếc loa 6 inch và một thanh đỡ luồng khí và chiết áp phản hồi trọng lực được đặt cách con thỏ khoảng 5 inch. Một sợi tơ được gắn vào một thanh được ghép cơ học với trục của chiết áp. Một chiếc móc kim loại nhỏ nối với đầu kia của sợi tơ được gắn vào một vòng nylon được khâu vào màng mí mắt phải của con thỏ. Tín hiệu từ chiết áp được khuếch đại và ghi lại bằng đồ họa. Lỗ của luồng khí được điều chỉnh để cung cấp một luồng nitơ nén 80 mm trong thời gian 100 mili giây từ vị trí cách vùng lưng của giác mạc bên phải khoảng 1/2 inch. Kích thích có điều kiện (CS) bao gồm âm 800 cy/giây 72 db SPL, được trình bày trong 600 mili giây.
Mười tám con thỏ bạch tạng, từ 85 đến 100 ngày tuổi, được phân vào một trong hai nhóm trong 2 ngày thích nghi, 8 ngày huấn luyện tiếp thu và 8 ngày giải trừ. Trong mỗi ngày tiếp nhận, một nhóm đối chứng gồm sáu con thỏ (nhóm C) nhận được trình bày ngẫu nhiên gồm 70 thử nghiệm riêng lẻ CS và 70 thử nghiệm kích thích không điều kiện (UCS) được giới hạn trong hai khối thử nghiệm ở các khoảng thời gian giữa các thử nghiệm ngẫu nhiên là 5, 10 và 15 giây ( trung bình là 10 giây). Một nhóm điều kiện hóa cổ điển gồm 12 con thỏ (nhóm E) đã nhận được 70 trình bày theo cặp về CS và UCS mỗi ngày với khoảng thời gian CS-UCS là 500 mili giây và các khoảng thời gian ngẫu nhiên giữa các thử nghiệm là 15, 20 và 25 giây (trung bình là 20 giây). Trong mỗi 2 ngày thích ứng, người ta đã đo được chuyển động màng tự phát ở cả hai nhóm bằng cách ghi lại tần suất phản hồi trong các khoảng thời gian tương ứng với 70 thử nghiệm CS-UCS được sử dụng để thu thập dữ liệu cho nhóm E.
Trong quá trình đáp ứng và tiếp nhận, tất cả chuyển động mở rộng màng có độ lệch ít nhất 1 mm so với đường cơ sở được ghi lại từ 0 đến 525 mili giây sau khi bắt đầu thử nghiệm. Khi giải trừ, khoảng thời gian được kéo dài đến 600 mili giây. Sự phân bố độ trễ phản hồi cho nhóm E trong quá trình tiếp nhận và giải trừ được hiển thị trong Hình 1.  Phía bên trái của hình cho thấy các phân phối đơn điệu và độ trễ có phương thức giảm một cách có hệ thống từ 525 mili giây vào ngày tiếp nhận thứ hai xuống còn 150 mili giây vào ngày thứ tám. Phát hiện này phù hợp với sự giảm dần về độ trễ phản ứng được báo cáo bởi Pavlov (4) đối với việc điều kiện hóa đồng thời phản ứng nước bọt. Một phân tích về phương sai của độ trễ trung bình của các phản ứng có điều kiện qua nhiều ngày cho thấy rằng độ trễ phản hồi giảm đáng kể (P< 0,01).
Phía bên trái của hình cho thấy các phân phối đơn điệu và độ trễ có phương thức giảm một cách có hệ thống từ 525 mili giây vào ngày tiếp nhận thứ hai xuống còn 150 mili giây vào ngày thứ tám. Phát hiện này phù hợp với sự giảm dần về độ trễ phản ứng được báo cáo bởi Pavlov (4) đối với việc điều kiện hóa đồng thời phản ứng nước bọt. Một phân tích về phương sai của độ trễ trung bình của các phản ứng có điều kiện qua nhiều ngày cho thấy rằng độ trễ phản hồi giảm đáng kể (P< 0,01).
Trong giải trừ, hai chế độ xuất hiện trong các bản phân phối. Mặc dù tần suất phản ứng giảm dần theo những ngày giải trừ, nhưng dường như không có sự thay đổi mang tính hệ thống nào về độ trễ có phương thức. Phương thức đầu tiên ở bên trái chủ yếu là chức năng đáp ứng sơ cấp ở hai con thỏ. Việc kiểm tra sự phân bố độ trễ của các phản hồi ở cả hai nhóm trong quá trình thích ứng (không hiển thị) và nhóm C trong tiếp nhận và giải trừ (không hiển thị) cho thấy chúng không có hệ thống và không thường xuyên và về cơ bản giống như được hiển thị trong Hình 1 của nhóm E ở ngày đầu tiên tiếp nhận. Do đó, không có bằng chứng nào trong dữ liệu về phản ứng phản xạ đối với CS hoặc về sự nhạy cảm.
Tỷ lệ phần trăm phản hồi cho cả hai nhóm trong việc thích ứng, tiếp nhận và giải trừ được thể hiện trong Hình 2.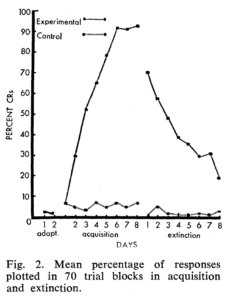 Để thích ứng, tỷ lệ phản hồi tự phát là khoảng 1,5 cho cả hai nhóm. Trong quá trình tiếp nhận, phản hồi dành cho nhóm C không tăng theo ngày và không bao giờ vượt quá mức 6%. Nhóm E cho thấy số phản hồi tăng đều đặn từ 6% trong ngày đầu tiên lên mức tiệm cận khoảng 92% trong 3 ngày tiếp nhận cuối cùng.
Để thích ứng, tỷ lệ phản hồi tự phát là khoảng 1,5 cho cả hai nhóm. Trong quá trình tiếp nhận, phản hồi dành cho nhóm C không tăng theo ngày và không bao giờ vượt quá mức 6%. Nhóm E cho thấy số phản hồi tăng đều đặn từ 6% trong ngày đầu tiên lên mức tiệm cận khoảng 92% trong 3 ngày tiếp nhận cuối cùng.
Tuy nhiên, trong nghiên cứu được báo cáo trước đây về việc điều kiện hóa mí mắt bên ngoài (2), mức độ điều kiện hóa cao nhất đạt được là 72%. So sánh i-test về tỷ lệ phần trăm trung bình của các phản ứng có điều kiện của nhóm E vào ngày đầu tiên và ngày thứ tám thu được là rất có ý nghĩa (P= .001). Nhóm H cho thấy khả năng chống giải trừ đáng kể. Phản hồi trung bình là từ 71% vào ngày đầu tiên đến 20% vào ngày thứ tám. Mặt khác, ở nhóm C sự giải trừ và sự thích ứng gần như song song (5).



Đăng nhập để bình luận.