
Mạch thần kinh và cơ chế khả biến duy trì phản xạ điều kiện chớp mắt



- Bs Đỗ Thị Thúy Anh
Phản xạ có điều kiện chớp mắt Pavlovian đã được sử dụng rộng rãi như một hệ thống mô hình để nghiên cứu các cơ chế thần kinh làm cơ sở cho việc học tập kết hợp. Việc trì hoãn phản xạ chớp mắt phụ thuộc vào tiểu não trung gian cùng bên với mắt có phản xạ điều kiện. Bằng chứng ủng hộ mô hình khả biến hai vị trí trong tiểu não với sự ức chế dài hạn (long-term depression LTD) của các synap sợi song song/parallel fiber trên tế bào Purkinje và điện thế hóa dài hạn (long-term potentiation LTP) của các synap sợi rêu/mossy fiber trên neuron ở nhân xen trước/ anterior interpositus nucleus. Đầu vào kích thích có điều kiện và kích thích không điều kiện phát sinh từ nhân cầu não và nhân olive dưới, hội tụ ở vỏ tiểu não và nhân sâu. Các truyền dẫn từ nhân cảm giác dưới vỏ não đến nhân cầu não cần thiết cho phản xạ điều kiện chớp mắt đang bắt đầu được xác định, và các nghiên cứu gần đây chỉ ra rằng có những tương tác động giữa nhân đồi thị cảm giác và tiểu não trong quá trình điều kiện hóa chớp mắt. Tín hiệu từ tiểu não được truyền tới tế bào nhân đỏ và sau đó tới các nhân vận động tạo ra (các) phản ứng chớp mắt. Tiến bộ to lớn đã được thực hiện trong việc xác định các cơ chế thần kinh của việc trì hoãn phản xạ điều kiện chớp mắt nhưng vẫn còn những lỗ hổng đáng kể trong sự hiểu biết của chúng ta về các cơ chế khả biến và mạch thần kinh cần thiết trong quá trình học tập của tiểu não.
Giới thiệu
Phản xạ điều kiện chớp mắt là một mô hình học tập kết hợp lần đầu tiên được phát triển để sử dụng cho những thành phần tham gia là con người vào những năm 1920 (Cason 1922). Ban đầu nó được đánh giá là một phương pháp nghiên cứu khả năng học tập và chức năng hệ thần kinh cấp cao mà không bị nhiễu thông tin từ lời nói, sự xem xét nội tâm hoặc kinh nghiệm trước đây trí nhớ) với các kết hợp tương tự. Quy trình này bao gồm việc đưa ra một kích thích có điều kiện (CS), điển hình là âm thanh hoặc ánh sáng, được kết hợp với một kích thích không điều kiện (US) có tác dụng kích thích mí mắt đóng lại một cách đáng tin cậy, chẳng hạn như một luồng khí hoặc kích thích điện ngắn gần mắt. Con người thường sẽ thể hiện phản ứng không điều kiện (alpha) biên độ thấp có độ trễ ngắn đối với CS thính giác. Sau nhiều lần thử nghiệm CS-US, việc nhắm mắt có điều kiện (phản ứng có điều kiện [CR]) xảy ra trong CS. Việc đóng mí mắt tối đa đối với CR thường xảy ra gần ngay sau thời điểm khởi phát US. Sau đó, một số thiếu sót của mô hình đã được xác định, đáng chú ý nhất là sự hiện diện của phản ứng alpha và đáp ứng tự nguyện ở những thành phần tham gia là con người, những người tỉnh thức (aware) rõ ràng về khả năng kích thích ngẫu nhiên. Những thiếu sót này và nhu cầu về mô hình động vật cho nghiên cứu khoa học thần kinh xâm lấn đã dẫn đến sự phát triển của mô hình màng mí mắt/ nictitating membrane và chớp mắt của thỏ (Gormezano và cộng sự 1962; Schneiderman và cộng sự 1962; Gormezano 1966). Thỏ chịu đựng được sự kiềm chế tốt, không biểu hiện phản ứng alpha và có thể dễ dàng thực hiện các biện pháp chính xác về việc nhắm mắt và chuyển động màng mí mắt (Gormezano 1966). Hầu hết công việc ban đầu về cơ chế thần kinh làm cơ sở cho phản xạ điều kiện chớp mắt được thực hiện trên thỏ, nhưng mô hình này cũng đã được áp dụng cho ếch, rùa, chuột nhắt, chồn sương, cừu, chó, khỉ và mèo. Mối lo ngại khi sử dụng các loài không phải thỏ là hầu hết chúng sẽ hiển thị phản ứng alpha nếu các thông số CS và US không được điều chỉnh hợp lý. Ví dụ: chúng tôi đã phát hiện ra rằng phản ứng alpha ở loài gặm nhấm có thể được loại bỏ gần như hoàn toàn bằng cách giảm tần số (2 kHz) và biên độ (70 dB đối với chuột, 80 – 85 dB đối với chuột) của CS thính giác và thời gian US (25 mili giây). Mặc dù có một tài liệu phong phú về hành vi từ các nghiên cứu về phản xạ điều kiện chớp mắt ở người và động vật thí nghiệm, đánh giá này sẽ tập trung chủ yếu vào những phát hiện sinh học thần kinh từ các nghiên cứu về duy trì phản xạ điều kiện.
Nháy mắt là gì?
Điều quan trọng cần lưu ý ở điểm này là phản ứng không điều kiện (UR) và CR trong điều kiện hóa chớp mắt khác nhau và khác với chớp mắt tự phát về chuyển động học và thần kinh điều khiển (Gruart et al. 1995, 2000; Trigo et al. 1999; Schade Powers và cộng sự 2010). Các đặc tính nhịp của CR được xác định bởi khoảng thời gian giữa các lần kích thích, với đỉnh CR gần thời điểm khởi phát của US, trong khi các đặc tính nhịp của UR được xác định bởi các thông số của US (Gormezano và cộng sự 1983). Hơn nữa, các kiểu chớp mắt (UR) khác nhau được tạo ra bởi các kích thích khác nhau (Gruart và cộng sự 1995; Trigo và cộng sự 1999); lĩnh vực này thường bỏ qua những khác biệt như vậy, có lẽ gây bất lợi cho chúng ta. “Nháy mắt” được sử dụng trong bài nói về chuyển động của mí mắt và màng mí mắt bởi vì, như được thảo luận dưới đây, mạch thần kinh tiền vận động bên dưới các CR tương ứng là giống nhau. Tuy nhiên, các nhân vận động tạo ra chuyển động của mí mắt và màng mí mắt là khác nhau (xem phần Mạch Đầu ra Phản hồi). Trọng tâm của bài tổng quan này là về các cơ chế điều kiện hóa tiền vận động và do đó sẽ sử dụng thuật ngữ “nháy mắt” để chỉ các chuyển động của mí mắt và màng mí mắt, cũng như CR và UR.
Vai trò thiết yếu của tiểu não trong duy trì phản xạ điều kiện
Brogden và Gantt (1942) là một trong những người đầu tiên chứng minh vai trò của tiểu não trong việc học tập kết hợp bằng cách điều kiện hóa các chuyển động rời rạc khi sử dụng kích thích điện của tiểu não khi US kết hợp với CS thính giác. Eccles và cộng sự. (1967) sau đó đưa ra giả thuyết rằng tiểu não đóng vai trò trong việc học kỹ năng, dựa trên sinh lý học và giải phẫu vỏ não tiểu não. Giả thuyết này được chính thức hóa trong các mô hình tính toán của Marr (1969) và Albus (1971). Một thành phần quan trọng của mô hình Albus là các tế bào Purkinje ở tiểu não trải qua quá trình ức chế liên quan đến học tập. Giải thích hiện nay về cơ chế này là sự ức chế tế bào Purkinje giải phóng các nhân sâu khỏi sự ức chế trương lực của nó. Cơ chế ức chế hóa dài hạn (LTD) này đã được xác minh bằng nhiều sản phẩm in vivo và in vitro khác nhau (Ito và Kano 1982; Linden và Connor 1991, 1995; Linden và cộng sự 1991; Linden 1994). Một trong những nghiên cứu đầu tiên cho thấy mối tương quan giữa tiểu não với khả năng học tập là thí nghiệm của Gilbert và Thach (Gilbert và Thach 1977), họ nhận thấy giảm hoạt động nhọn sóng đơn giản của tế bào Purkinje, phù hợp với giả thuyết LTD, trong một nhiệm vụ yêu cầu khỉ sửa đổi chuyển động cổ tay để bù đắp đối với những thay đổi về mang vác (load). Họ cũng tìm thấy bằng chứng cho thấy sự gia tăng hoạt động nhọn sóng phức tạp tương ứng với những thay đổi mới khi mang vác. Nghiên cứu của Gilbert và Thach cho thấy mối tương quan đáng chú ý giữa việc học và việc sửa lỗi của một chuyển động đơn giản nhưng không chứng minh được rằng các tế bào Purkinje liên quan đến nhiệm vụ cần thiết cho việc học.
Thompson và các đồng nghiệp là những người đầu tiên chứng minh rằng tiểu não đóng vai trò thiết yếu trong việc điều kiện hóa chớp mắt (McCormick và cộng sự 1981, 1982; Lincoln và cộng sự 1982). Trong các nghiên cứu ban đầu của họ, các tổn thương do hủy bỏ ở bán cầu tiểu não cùng bên với mắt điều kiện hóa đã được phát hiện là có thể loại bỏ CR trong khi vẫn giữ nguyên khả năng điều điều kiện hóa của mắt đối diện. Các tổn thương bao gồm nhân sâu cùng bên là ảnh hưởng nhất (xem thêm Yeo và cộng sự 1985a). Đáng chú ý, các tổn thương không ảnh hưởng đến UR, cho thấy hiệu suất của phản ứng không bị ảnh hưởng bởi các tổn thương loại bỏ CR. Các tổn thương chính xác hơn, bao gồm các tổn thương độc thần kinh không phải-sợi, ở nhân tiểu não, đã xác định phần phía ngoài của nhân xen/ trước là quần thể tế bào thần kinh cần thiết cho việc duy trì (Lavond et al. 1985). Hơn nữa, các tổn thương của nhân xen/ interpositus sẽ ngăn chặn vĩnh viễn quá trình tiếp nhận (Steinmetz và cộng sự 1992a,b) và loại bỏ khả năng lưu giữ sau khi củng cố trí nhớ (Christian và Thompson 2005). Việc vô hiệu hóa nhân xen trước bằng cách làm lạnh, lidocain hoặc muscimol sẽ loại bỏ CR là có thể đảo ngược (Chapman và cộng sự 1990; Clark và cộng sự 1992; Krupa và cộng sự 1993; Hardiman và cộng sự 1996; Garcia và Mauk 1998; Bao và cộng sự. 2002; Aksenov và cộng sự 2004; Freeman và cộng sự 2005a). Một số báo cáo đã thách thức những phát hiện và cách giải thích của các nghiên cứu được liệt kê ở trên (Welsh và Harvey 1989, 1991; Jimenez-Diaz et al. 2004). Tuy nhiên, hầu hết các phát hiện từ các nghiên cứu về tổn thương và bất hoạt đều chỉ ra rằng nhân xen trước là cần thiết để tiếp nhận và lưu giữ CR.
Các nghiên cứu sinh lý thần kinh chứng minh thêm rằng các neuron ở nhân xen trước, chứ không phải phía sau, biểu hiện một bùng nổ truyền dẫn ở động vật đã học tập, tạo mô hình biên độ/thời gian của CR (Hình 1; McCormick và cộng sự 1982; McCormick và Thompson 1984; Berthier và Moore 1990; Gould và Steinmetz 1996; Freeman và Nicholson 2000; Nicholson và Freeman 2002; Choi và Moore 2003; Halverson và cộng sự 2010). Hơn nữa, bùng nổ hoạt động ở các neuron nhân xen trước này chỉ được quan sát thấy khi CR xảy ra và dẫn trước các thử nghiệm-nội bộ CR (Nicholson và Freeman 2002; Choi và Moore 2003; Green và Arenos 2007; Halverson và cộng sự 2010) . Cấu hình hoạt động của các neuron nhân xen trước cho thấy rằng các neuron này đang điều khiển các neuron vận động tạo ra CR. Bằng chứng sâu hơn ủng hộ mối quan hệ nhân quả giữa hoạt động của neuron nhân xen trước và sản xuất CR đến từ các nghiên cứu sử dụng kích thích điện thông qua các điện cực ghi để tạo ra sự đóng mí mắt (McCormick và Thompson 1984; Freeman và Nicholson 2000; Halverson và cộng sự 2010). Trên thực tế, việc đóng mí mắt có thể được tạo ra bằng cách kích thích nhân xen trước ở trước khi tập luyện, cho thấy rằng có một mối liên hệ chặt chẽ giữa tiểu não và các neuron tiền vận động tạo ra CR. Các tổn thương và nghiên cứu ghi ngoại bào cung cấp bằng chứng thuyết phục rằng hoạt động ở nhân xen trước thúc đẩy sản xuất CR.
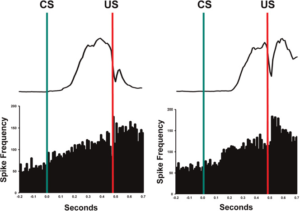 Hình 1. Ví dụ về hoạt động đơn vị (nhọn sóng/giây) được ghi lại từ nhân xen trước và hoạt động EMG mí mắt trung bình tương ứng ở những con chuột được huấn luyện tốt trong các thử nghiệm CS-US. Đường màu xanh lá cây tượng trưng cho sự khởi đầu của CS và đường màu đỏ tượng trưng cho sự khởi đầu của US. Các thanh trong biểu đồ là 10 mili giây. Lưu ý sự gia tăng hoạt động sau khi bắt đầu CS.
Hình 1. Ví dụ về hoạt động đơn vị (nhọn sóng/giây) được ghi lại từ nhân xen trước và hoạt động EMG mí mắt trung bình tương ứng ở những con chuột được huấn luyện tốt trong các thử nghiệm CS-US. Đường màu xanh lá cây tượng trưng cho sự khởi đầu của CS và đường màu đỏ tượng trưng cho sự khởi đầu của US. Các thanh trong biểu đồ là 10 mili giây. Lưu ý sự gia tăng hoạt động sau khi bắt đầu CS.
Các tổn thương ở vỏ tiểu não thường dẫn đến suy giảm nghiêm trọng khả năng điều kiện hóa chớp mắt (Lavond và Steinmetz 1989; Garcia et al. 1999). Trong một số trường hợp, việc học tập ở động vật bị tổn thương vỏ tiểu não cuối cùng cũng xảy ra với sự huấn luyện nhắm (Lavond và Steinmetz 1989; Chen và cộng sự 1996, 1999; Nolan và Freeman 2006). Ví dụ, Lavond và Steinmetz (1989) đã tạo ra các tổn thương do hủy bỏ lớn ở vỏ tiểu não bao gồm gần như toàn bộ bán cầu ở thỏ và phát hiện ra rằng một số con thỏ này có được khả năng điều kiện hóa chớp mắt khi được huấn luyện nhắm. Những minh chứng đặc biệt thuyết phục về vai trò của vỏ tiểu não trong việc thu nhận khả năng điều kiện hóa chớp mắt đến từ các nghiên cứu kiểm tra khả năng điều kiện hóa sau khi toàn bộ tế bào Purkinje bị mất đi. Các tế bào Purkinje được cho là có tính khả biến trong vỏ tiểu não trong quá trình học tập và các sợi trục của chúng tạo thành toàn bộ đầu ra của vỏ tiểu não. Do đó, sự cạn kiệt của các tế bào Purkinje sẽ loại bỏ ảnh hưởng của độ khả biến vỏ tiểu não và sự ức chế trương lực lên nhân sâu. Những con chuột bị đột biến dẫn đến mất tế bào Purkinje toàn thể cho thấy khả năng tiếp thu chậm hơn và tiệm cận thấp hơn (Chen và cộng sự 1996, 1999). Hơn nữa, những con chuột bị mất toàn bộ tế bào Purkinje do sử dụng chất độc miễn dịch cũng cho thấy khả năng điều kiện hóa chớp mắt diễn ra rất chậm (Nolan và Freeman 2006). Ngược lại, Yeo và các đồng nghiệp phát hiện ra rằng những con thỏ được phong tỏa thụ thể AMPA không đạt được khả năng điều kiện hóa chớp mắt (Attwell và cộng sự 2001). Tuy nhiên, một thiếu sót của nghiên cứu này là thỏ chỉ được huấn luyện trong 200 thử nghiệm với việc phong tỏa thụ thể AMPA. Vì vậy, có thể những con thỏ này cuối cùng đã học được khi được huấn luyện thêm trong quá trình phong tỏa AMPA. Những phát hiện từ các nghiên cứu sử dụng nhiều loại tổn thương khác nhau nhìn chung đã chỉ ra rằng vỏ tiểu não đóng vai trò quan trọng trong việc tiếp nhận điều kiện hóa của chớp mắt, nhưng việc học tập có thể chậm chạp nếu không có vỏ đại não.
Những phát hiện từ các nghiên cứu kiểm tra ảnh hưởng của tổn thương vỏ não đến khả năng duy trì khả năng điều kiện hóa chớp mắt là khá khác nhau. Hủy lớn vỏ tiểu não của bán cầu dẫn đến mất CR ban đầu và việc học lại một phần hoặc toàn bộ sau đó bằng việc đào tạo lại (Lavond và cộng sự 1987; Yeo và Hardiman 1992; Harvey và cộng sự 1993; Woodruff-Pak và cộng sự 1993). Một kết quả tương tự được tìm thấy với tình trạng mất tế bào Purkinje do độc tố miễn dịch gây ra trên khắp vỏ tiểu não (Nolan và Freeman 2005). Trong một số nghiên cứu, mức độ phục hồi phụ thuộc vào mức độ tổn thương của thùy H VI và các khu vực lân cận của thùy ansiform (Yeo và Hardiman 1992; Harvey và cộng sự 1993). Mức độ tập luyện trước khi bị tổn thương cũng có thể liên quan đến tốc độ và mức độ hồi phục sau tổn thương đi sau tổn thương tiểu thùy H VI (Harvey et al. 1993). Các tổn thương nhỏ hơn của vỏ tiểu não khu trú ở thùy trước gây ra sự gián đoạn thời gian CR mà không làm giảm đáng kể tỷ lệ CR (Perrett et al. 1993). Do đó, có thể thùy trước và thùy HVI có những đóng góp khác nhau trong việc điều kiện hóa chớp mắt sau khi học.
Các nghiên cứu dược lý về sự tương tác giữa vỏ tiểu não và nhân sâu đã tiết lộ nhiều hơn về vai trò tương ứng của chúng trong cơ chế điều hòa và lưu trữ trí nhớ so với các nghiên cứu về tổn thương. Những tranh cãi về vô số yếu tố có thể dẫn đến các kết quả khác nhau sau tổn thương đã dẫn đến các nghiên cứu sử dụng phương pháp tách vỏ tiểu não khỏi nhân sâu bằng thuốc (Mauk 1997). Đây là một cách tiếp cận có thể thực hiện được bằng thực nghiệm vì toàn bộ đầu ra của vỏ tiểu não đi đến nhân tiểu não và tiền đình, và đó là GABAergic. Do đó, các loại thuốc ngăn chặn sự truyền dẫn qua khớp thần kinh GABAergic sẽ chặn đầu vào vỏ não đến các nhân sâu. Một số nghiên cứu đã sử dụng phương pháp ngắt kết nối dược lý này và phát hiện ra rằng việc ngăn chặn sự truyền qua khớp thần kinh GABA-A bằng picrotoxin ức chế kênh clorua GABA-A hoặc gabazine đối kháng thụ thể GABA-A dẫn đến CR có độ trễ ngắn; nghĩa là, thời gian CR đã bị gián đoạn (Garcia và Mauk 1998; Medina và cộng sự 2001; Ohyama và cộng sự 2003, 2006; Parker và cộng sự 2009; Vogel và cộng sự 2009). Hơn nữa, tỷ lệ phần trăm CR chỉ giảm khi dùng picrotoxin liều cao. Mối lo ngại về việc sử dụng picrotoxin để ngăn chặn sự truyền qua khớp thần kinh là nó cũng gây mất ức chế trương lực của các tế bào thần kinh nhân sâu (Aksenov và cộng sự 2004). Một nghiên cứu của Thompson và các đồng nghiệp đã sử dụng lần lượt sử dụng muscimol và picrotoxin trong nhân xen để ngăn chặn đầu vào synap GABA-A từ vỏ tiểu não trong khi điều chỉnh sự ức chế trương lực trong nhân xen (Bao và cộng sự 2002). Khi quy trình tiêm truyền tuần tự này được sử dụng ở những con thỏ đã được điều kiện hóa chớp mắt, CR ban đầu bị loại bỏ nhưng được phục hồi khi truyền picrotoxin, cho thấy rằng việc giải phóng khỏi chất ức chế trương lực là đủ để khôi phục CR. Tuy nhiên, CR được phục hồi có độ trễ khởi phát ngắn hơn cho thấy thời gian CR bị gián đoạn do chặn quá trình truyền synap từ các tế bào Purkinje. Do đó, giống như trong các nghiên cứu trước đây, việc ngăn chặn sự truyền synap GABA-A từ tế bào Purkinje đến nhân xen làm giảm thời gian của CR. Những phát hiện này cũng chỉ ra rằng dấu vết trí nhớ đủ cho CR cơ bản, không xác định thời gian được thiết lập trong nhân xen trong quá trình điều kiện hóa chớp mắt (Ohyama et al. 2006).
Tại sao kết quả từ nghiên cứu tổn thương và nghiên cứu dược lý lại khác nhau về việc duy trì tần số CR? Các tổn thương thường có thời gian phục hồi theo sau >1 tuần. Các vùng não ly tâm và hướng tâm đến vị trí tổn thương có thể trải qua những thay đổi phản ứng và bù trừ sau tổn thương có thể ảnh hưởng đến biểu hiện sau phẫu thuật. Những thay đổi tạm thời về chức năng synap trong nhân sâu tạo ra những thay đổi về hoạt động trương lực và pha trong mạch xuôi dòng nhưng không tạo ra những thay đổi mãn tính trong mạch thần kinh được thấy ở các tổn thương. Các tổn thương vỏ tiểu não đã được chứng minh là làm tăng sự tăng sinh thần kinh đệm trong nhân xen (Yeo và cộng sự 1985b; Harvey và cộng sự 1993). Tác động của bệnh thần kinh đệm lên chức năng neuron nhân xen chưa được xác định, nhưng việc mất đi sự phân bố tế bào Purkinje của các nhân sâu có thể dẫn đến vô số thay đổi khác trong chức năng thần kinh. Ví dụ, neuron trong nhân xen có thể tạo ra những thay đổi bù trừ về hiệu quả của synap glutamatergic hoặc tính dễ bị kích thích nội tại sau khi vỏ não mất đi khả năng ức chế trương lực để ngăn ngừa độc tính kích thích; Ngược lại, những thay đổi bù trừ này có thể làm giảm hiệu suất của CR bằng cách làm suy yếu đầu vào CS từ các sợi rêu. Một cơ chế khác được đề xuất bởi các nghiên cứu cho thấy sự thoái hóa của nhân olive/trám dưới sau các tổn thương vỏ tiểu não (Yeo và cộng sự 1985b; Lavond và cộng sự 1987). Việc mất đi các neuron olive dưới và tổn thương vỏ não sẽ làm giảm khả năng kích hoạt sợi leo 2 bên của các nhân sâu, điều này có thể làm giảm khả năng duy trì tính khả biến trong các nhân sâu bằng cách loại bỏ một nguồn củng cố thần kinh. Các yếu tố nguyên nhân dẫn đến sự khác biệt giữa tác động của tổn thương vỏ tiểu não và sự mất kết nối dược lý chưa được xác định nhưng có thể là do tác động phản ứng mãn tính và cấp tính của các suy thoái sinh lý của neuron trong nhân xen.
Nghiên cứu của Gilbert và Thach (1977) cho thấy các ví dụ về tế bào Purkinje có biểu hiện giảm hoạt động nhọn sóng đơn giản trong quá trình thích ứng các chuyển động của cổ tay với mang vác/ load. Sự giảm hoạt động nhọn sóng đơn giản này được hiểu là phù hợp với lý thuyết Albus về khả năng học tập của tiểu não (Albus 1971). Một số nghiên cứu đã tìm thấy sự giảm tương tự trong hoạt động nhọn sóng đơn giản trong quá trình điều kiện hóa chớp mắt. Một tỷ lệ lớn tế bào Purkinje ở thùy trước và các phần lân cận của tiểu thùy H VI đã làm giảm hoạt động nhọn sóng đơn giản trong CS ở những con thỏ được huấn luyện tốt, đặc biệt là sau đó trong khoảng CS-US nơi CR được sản xuất (Green và Steinmetz 2005). Sự giảm hoạt động nhọn sóng đơn giản này cũng được định thời gian thích hợp cho các khoảng thời gian kích thích khác nhau. Sau đó, một nghiên cứu có phần hạn chế nhưng được kiểm soát tốt đã phát hiện ra rằng điều kiện hóa chớp mắt bằng cách sử dụng kích thích điện ở cẳng tay khi CS ở chồn sương bị mất não đã tạo ra sự tạm dừng hoạt động của tế bào Purkinje trong vùng chớp mắt của vỏ tiểu não ( Jirenhed và cộng sự 2007). Việc tạm dừng trong hoạt động nhọn sóng đơn giản đã bị bãi bỏ bằng quá trình huấn luyện giải trừ nhưng được tái đáp ứng lại bằng việc nối lại các cặp CS-US. Việc giải thích những phát hiện này phần nào bị hạn chế do không có phản ứng hành vi, loại trừ khả năng liên quan đến sự phát triển của các khoảng dừng trong hoạt động nhọn sóng đơn giản với sự phát triển của CR qua các thử nghiệm hoặc liên quan đến thời gian tạm dừng với thời gian của CR. Một nghiên cứu quan trọng cho lĩnh vực này là ghi lại hoạt động của các tế bào Purkinje liên quan đến chớp mắt trong quá trình tiếp nhận và giải trừ ở động vật còn nguyên vẹn để liên hệ các khoảng dừng trong các nhọn sóng đơn giản với sự phát triển và giải trừ của hành vi CR.
Rõ ràng là vỏ tiểu não và nhân xen trước cùng bên với mắt điều kiện hóa đóng vai trò quan trọng trong việc tiếp nhận và duy trì khả năng điều kiện hóa chớp mắt. Các tổn thương và bất hoạt của các vùng vỏ não có liên quan hoặc nhân xen trước làm suy giảm khả năng tiếp nhận và các neuron trong các vùng này cho thấy đặc điểm hoạt động tương quan với sự phát triển của CR chớp mắt trong các thử nghiệm CS-US và trong mỗi thử nghiệm. Hơn nữa, động lực của hoạt động thần kinh trong các thử nghiệm chỉ ra rằng việc tạm dừng hoạt động nhọn sóng đơn giản của tế bào Purkinje sẽ giải phóng nhân xen trước khỏi sự ức chế và bùng nổ hoạt động từ nhân sẽ thúc đẩy các neuron tiền vận động và vận động tạo ra CR.
Vai trò của tiểu não trong học tập kết hợp đã được kiểm tra, chủ yếu bằng cách sử dụng điều kiện hóa chớp mắt. Tuy nhiên, tiểu não rất cần thiết cho các CR kiểu Pavlov khác, bao gồm cả khả năng uốn cong chi trước có điều kiện ở mèo (Voneida 2000) và khả năng uốn cong chi sau có điều kiện ở thỏ (Mojtahedian và cộng sự 2007). Tiểu não cũng góp phần điều chỉnh các chuyển động đã học được (Gilbert và Thạch 1977), hiệu chỉnh tầm nhìn (Norris và cộng sự 2011), điều chỉnh phản xạ tiền đình-mắt (Raymond và cộng sự 1996; Boyden và cộng sự 2004), và học chuyển động mắt theo đuổi nhịp nhàng (Medina và Lisberger 2008).
Định vị dấu vết trí nhớ trong tiểu não
Các nghiên cứu được xem xét cho đến nay cho thấy rằng tiểu não cần thiết cho việc tiếp nhận và duy trì khả năng điều kiện hóa chớp mắt. Tuy nhiên, không có nghiên cứu nào trong số này chứng minh rõ ràng rằng trí nhớ bên dưới CR chỉ được lưu trữ trong tiểu não.
Sự bất hoạt có thể đảo ngược của các phần khác nhau của mạch tiểu não đã được sử dụng một cách có hệ thống để lập luận rằng bộ nhớ điều kiện hóa chớp mắt phải được lưu trữ trong tiểu não. Logic của phương pháp này là việc ngừng hoạt động của vùng não lưu trữ bộ nhớ sẽ không dẫn đến CR trong quá trình tiếp nhận và quan trọng hơn là sẽ không tiết kiệm được sau khi ngừng hoạt động. Nghĩa là, tốc độ tiếp nhận sau khi ngừng hoạt động phải giống như tỷ lệ tiếp nhận ban đầu trong nhóm đối chứng thích hợp. Ngược lại, nếu việc bất hoạt ngăn cản CR trong quá trình tiếp nhận, nhưng lại tiết kiệm được sau khi loại bỏ trạng thái bất hoạt, thì kết luận là vùng bất hoạt có liên quan đến biểu hiện học tập nhưng không lưu trữ bộ nhớ bên dưới CR. Việc ngừng hoạt động của nhân tiểu não và các vùng vỏ não lân cận sẽ ngăn chặn CR trong quá trình tiếp nhận và dẫn đến không tiết kiệm được thời gian luyện tập tiếp theo nếu không bất hoạt (Clark và cộng sự 1992; Krupa và cộng sự 1993; Krupa và Thompson 1997; Freeman và cộng sự 2005a). Những phát hiện này chỉ ra rằng các cấu trúc hướng tâm đến tiểu não không thể lưu trữ ký ức, nhưng một khả năng khác là các cấu trúc ly tâm có thể lưu trữ ký ức, và việc bất hoạt tiểu não chỉ đơn giản là chặn sự chuyển tiếp kích thích ly tâm đến vị trí lưu trữ ký ức ở cuối dòng. Việc bất hoạt đường đầu ra của tiểu não, cuống tiểu não trên, nhân đỏ hoặc nhân vận động cần thiết để tạo ra (các) phản ứng chớp mắt sẽ dẫn đến việc loại bỏ CR trong quá trình tiếp nhận nhưng hoàn toàn tiết kiệm được trong quá trình huấn luyện tiếp theo mà không bị bất hoạt (Clark và Lavond 1993; Krupa và cộng sự 1993, 1996; Krupa và Thompson 1995). Hơn nữa, việc bất hoạt nhân đỏ sẽ ngăn chặn CR nhưng không ảnh hưởng đến hoạt động liên quan đến học tập trong tiểu não (Chapman và cộng sự 1990). Những phát hiện này chỉ ra rằng nhân đỏ và nhân vận động lần lượt là các khu vực tiền vận động và vận động, nằm ở cuối vùng lưu trữ bộ nhớ.
Bất hoạt có thể đảo ngược cũng đã được sử dụng để kiểm tra sự củng cố của điều kiện hóa chớp mắt. Việc ngừng hoạt động của vỏ tiểu não sau khi huấn luyện sẽ ngăn chặn việc duy trì khả năng điều kiện hóa chớp mắt ở thỏ, cung cấp thêm bằng chứng cho thấy ký ức cơ bản về CR của chớp mắt được lưu trữ trong tiểu não (Attwell và cộng sự 2002; Cooke và cộng sự 2004). Thật không may, những nghiên cứu này có phần hạn chế vì việc bất hoạt sau huấn luyện chỉ được sử dụng trong 200 thử nghiệm, làm tăng khả năng thỏ có thể học được nếu được huấn luyện nhiều hơn. Tuy nhiên, những phát hiện này bổ sung thêm bằng chứng cho thấy trí nhớ làm cơ sở điều kiện hóa chớp mắt được lưu trữ trong tiểu não. Những phát hiện này đặc biệt gợi ý rằng có bộ nhớ lưu trữ bên trong vỏ não, như đề xuất của Albus (1971) và Marr (1969).
Một loạt thí nghiệm sau đó đã chứng minh bằng chứng về việc lưu trữ trí nhớ trong nhân xen trước (Ohyama và cộng sự 2006). Cách tiếp cận của nghiên cứu này là ngắt kết nối vỏ tiểu não khỏi nhân xen về mặt dược lý, dẫn đến CR có độ trễ ngắn, sau đó kiểm tra các cơ chế tạo ra những phản ứng kém thời gian này bằng cách sử dụng các thao tác dược lý và kích thích điện. Phân tích này loại trừ các cơ chế khả biến hướng tâm và ly tâm đến nhân xen trước. Kết quả chỉ ra rằng biểu hiện cơ bản của bộ nhớ của CR có độ trễ ngắn rất có thể được lưu trữ trong nhân xen trước.
Một kết luận chung được rút ra từ Ohyama và cộng sự (2006) và những nghiên cứu khác cho rằng các cơ chế khả biến xảy ra trong vỏ não để hỗ trợ cơ chế giống LTD và trong nhân xen trước để hỗ trợ cơ chế giống LTP. Các cơ chế tế bào và phân tử làm cơ sở cho trí nhớ tiểu não đã được kiểm tra bằng nhiều phương pháp. Các thao tác di truyền và dược lý làm suy yếu vỏ tiểu não LTD thường làm suy giảm khả năng điều kiện hóa chớp mắt (Aiba và cộng sự 1994; Shibuki và cộng sự 1996; Ichise và cộng sự 2000; Kishimoto và cộng sự 2002; Lee và cộng sự 2009). Ví dụ, các thao tác di truyền của chức năng mGluR1 làm suy yếu LTD và điều kiện hóa chớp mắt ở chuột (Aiba và cộng sự 1994; Ichise và cộng sự 2000; Kishimoto và cộng sự 2002). Nhiều thao tác tổng thể hơn về cơ chế khả biến trong tiểu não, bao gồm phong tỏa tổng hợp protein, thụ thể NMDA và hoạt động kinase, tất cả đều làm suy yếu khả năng điều kiện hóa chớp mắt (Bracha và cộng sự 1998; Gomi và cộng sự 1999; Chen và Steinmetz 2000a, b). Một số nghiên cứu cũng đã tìm thấy bằng chứng về tính linh hoạt của cấu trúc trong tiểu não sau quá trình điều hòa chớp mắt. Ví dụ, số lượng synap bị kích thích ở trên mỗi neuron trong nhân xen trước đã tăng lên đáng kể sau khi điều kiện hóa chớp mắt ở những con chuột được huấn luyện tốt (Kleim và cộng sự 2002). Những thay đổi về hình thái synap nhưng không quan sát thấy số lượng trong nhân xen trước ở thỏ sau quá trình huấn luyện ít nhắm hơn (Weeks và cộng sự 2007). Những thay đổi trong cấu trúc synap cũng đã được tìm thấy ở vỏ tiểu não sau quá trình điều kiện hóa chớp mắt. Thỏ được điều kiện hóa chớp mắt trong 3 ngày cho thấy giảm các synap sợi song song có tính kích thích với tế bào Purkinje (Connor và cộng sự 2009). Do đó, các nghiên cứu hình thái về độ khả biến của synap đã tìm thấy những thay đổi ở synap có thể tạo ra cơ chế giống LTP ở nhân xen trước và cơ chế giống LTD ở vỏ tiểu não. Các nghiên cứu về cơ chế khả biến làm nền tảng cho việc điều kiện hóa chớp mắt nói chung cung cấp thêm bằng chứng về việc lưu trữ trí nhớ trong tiểu não.
Mạch đầu ra phản hồi
Các nghiên cứu sinh lý thần kinh được trích dẫn ở trên chỉ ra rằng hoạt động của nhân xen trước thúc đẩy CR gián tiếp thông qua các nueron tiền vận động. Vùng tế bào lớn/magnocellular của nhân đỏ nhận các truyền dẫn từ nhân tiểu não và dẫn tới các neuron vận động kiểm soát (các) phản ứng chớp mắt (Desmond và cộng sự 1983; Rosenfield và Moore 1983, 1985; Rosenfield và cộng sự 1985; Clark và Lavond 1993). Kích thích nhân đỏ, với các thông số thích hợp, tạo ra được phản ứng chớp mắt (Chapman và cộng sự 1988; Nowak và cộng sự 1997). Các neuron trong nhân đỏ phát triển hoạt động liên quan đến học tập gần giống với hoạt động nhân xen tạo dựng (Desmond và Moore 1991). Hơn nữa, sự bất hoạt của nhân đỏ sẽ loại bỏ CR trong khi vẫn giữ nguyên hoạt động liên quan đến học tập trong nhân xen (Chapman và cộng sự 1990). Phát hiện này và những phát hiện từ các nghiên cứu về sự bất hoạt (xem Bản địa hóa dấu vết trí nhớ trong phần Tiểu não) chỉ ra rằng nhân đỏ cần thiết để sản xuất CR nhưng không phải là nơi lưu trữ trí nhớ.
Neuron tế bào lớn nhân đỏ có các truyền dẫn kích thích tới các nhân vận động tạo ra các phản ứng chớp mắt khác nhau, bao gồm màng mí mắt và chuyển động mí mắt CR và UR. Chuyển động của màng mí mắt là một phản ứng thụ động đối với sự co lại của nhãn cầu được tạo ra bởi các neuron vận động trong nhân vận nhãn ngoài (nhân dây 6) và nhân vận nhãn ngoài phụ (Harvey và Gormezano 1981; Harvey và cộng sự 1984; Marek và cộng sự 1984; Disterhoft và cộng sự 1985; Weiss và Disterhoft 1985). Nhân vận nhãn ngoài phụ nhận đầu vào chuyển tiếp cảm giác từ nhân sinh ba và hệ lưới thân não (Harvey et al. 1984). Hoạt động của các neuron vận động vận nhãn ngoài có mô hình biên độ/thời gian phản ánh chặt chẽ phản ứng màng mí mắt ở thỏ đối với UR và CR (Cegavske và Thompson 1976; Cegavske và cộng sự 1979).
Chuyển động của mí mắt được tạo ra bởi nhân vận động mặt (Pellegrini và cộng sự 1995; Trigo và cộng sự 1999). Các nghiên cứu sinh lý thần kinh của Delgado-Garcia và các đồng nghiệp đã chứng minh các mô hình kích hoạt neuron vận động mặt dẫn đến mô hình động của việc nhắm mắt đối với các loại phản ứng khác nhau như thế nào (Trigo et al. 1999). Các cảm giác chuyển tiếp cần thiết để kích hoạt các neuron vận động chớp mắt bao gồm nhân sinh ba và tủy sống cổ (Pellegrini et al. 1995). Cả chuyển động của mí mắt và màng mí mắt UR đều được bắt đầu bởi các sợi cảm giác hướng tâm phát sinh từ giác mạc, trong trường hợp kích thích bằng luồng khí, hoặc từ vùng quanh ổ mắt, trong trường hợp kích thích sốc (điện?).
Mạch đầu vào kích thích không điều kiện
Các vùng sinh ba và tủy sống nhận cảm giác hướng tâm từ giác mạc và mặt truyền dẫn tới olive dưới. Các neuron ở phần lưng olive dưới phụ (dorsal accessory inferior olive DAO) thường tạo ra một hoặc hai điện thế hoạt động để phản ứng với US (Kim và cộng sự 1998; Nicholson và Freeman 2003a). Các neuron olive dưới hoạt động đồng bộ nhờ sự ghép cặp điện từ (Llinas và cộng sự 1974; Llinas và Yarom 1981a,b; Lang và cộng sự 1996). Các sợi trục của các neuron olive dưới trở thành các sợi leo trong tiểu não và có tạo sunap phụ với các neuron ở nhân sâu và tế bào Purkinje ở vỏ não (Sugihara và cộng sự 1999, 2001; Shinoda và cộng sự 2000). Các sợi leo tạo ra các tiếp xúc synap rất mạnh với các tế bào Purkinje, dẫn đến nhọn sóng phức hợp nhiều pha (Llinas và Sugimori 1980). Như đã đề cập ở trên, các mô hình ban đầu về chức năng tiểu não đã thừa nhận rằng các xung phức tạp cung cấp đầu vào từ US hoặc tín hiệu lỗi thúc đẩy sự biến đổi synap của tế bào Purkinje trong quá trình học tập (Marr 1969; Albus 1971).
Các tổn thương của olive dưới làm suy giảm khả năng điều hòa nhãn cầu (Mintz và cộng sự 1994; Welsh và Harvey 1998). Các tổn thương hoặc bất hoạt sau huấn luyện tạo ra sự giảm CR giống như giải trừ hoặc mất CR ngay lập tức (McCormick và cộng sự 1985; Yeo và cộng sự 1986; Welsh và Harvey 1998; Zbarska và cộng sự 2007). Sự giảm CR giống như bị giải trừ cũng được quan sát thấy sau các tổn thương ở olive dưới ở mèo (Voneida và cộng sự 1990). Cơ sở cho những phát hiện khác nhau về các tổn thương sau tập luyện ở olive dưới có phần bí ẩn vào thời điểm này, nhưng nó có thể liên quan đến quá trình phản ứng theo thời gian của tế bào Purkinje đối với việc mất đi sợi leo vào, tạo ra sự tạm dừng trong hoạt động nhọn sóng đơn giản, từ đó ảnh hưởng đến tính dễ bị kích thích của neuron trong nhân sâu (Bengtsson và cộng sự 2004). Có lẽ bằng chứng mạnh mẽ nhất cho thấy olive dưới là một phần của con đường US là việc kích thích điện của DAO có thể đóng vai trò là một US hiệu quả và hỗ trợ điều kiện hóa khi kết hợp với CS (Mauk và cộng sự 1986; Steinmetz và cộng sự 1989). Điều kiện hóa được thiết lập bằng kích thích DAO yêu cầu ghép nối CS-US với khoảng thời gian kích thích xen kẽ hỗ trợ điều kiện hóa với US ngoại vi. CR được thành lập với sự kích thích DAO vì US cũng có biểu hiện giải trừ trong các thử nghiệm chỉ dùng CS.
Olive dưới nhận được truyền dẫn phản hồi ức chế từ nhân tiểu não đóng vai trò quan trọng trong việc điều kiện hóa chớp mắt (Andersson và cộng sự 1988; Sears và Steinmetz 1991; Medina và cộng sự 2002; Nicholson và Freeman 2003a; Bengtsson và cộng sự 2004, 2007; Bengtsson và Hesslow 2006, Svensson và cộng sự 2006). Việc ngăn chặn chức năng GABAergic trong olive dưới tác động lên cơ chế ghép cặp điện tử, làm thay đổi tính đồng bộ của quá trình kích hoạt giữa các neuron olive và tăng tốc độ truyền dẫn cơ bản (Lang và cộng sự 1996; Medina và cộng sự 2002; Nicholson và Freeman 2003a, b). Phản hồi ức chế được cho là điều chỉnh độ khả biến của tiểu não bằng cách thay đổi tốc độ hoạt động của sợi ở vỏ não—tăng dẫn đến LTD và giảm dẫn đến LTP (Medina và cộng sự 2002). Phản hồi ức chế tiểu não đối với olive dưới cũng là cơ sở cho tác dụng ngăn chặn trong đó việc bổ sung một CS mới và dư thừa thì không tạo điều kiện hóa (Kim và cộng sự 1998).
Mạch đầu vào kích thích có điều kiện
Cuống tiểu não giữa bao gồm các sợi trục sợi rêu (sợi Mossy) chiếu vào tiểu não từ nhiều nguồn dưới vỏ khác nhau bao gồm nhân cầu não nền/basilar pontine nuclei (Shinoda et al. 1992, 2000; Mihailoff 1993). Nhân cầu não (PN) nhận được một lượng lớn thông tin cảm giác từ các vùng cảm giác vỏ đại não và nhân cảm giác dưới vỏ não (Glickstein và cộng sự 1972, 1980; Mihailoff và Watt 1981; Mihailoff và cộng sự 1985, 1989; Kosinski và cộng sự 1988; Legg và cộng sự 1989, Wells và cộng sự 1989). Phần lớn thông tin cảm giác này được chuyển tiếp đến nhân sâu tiểu não và lớp tế bào hạt của vỏ não (Shinoda et al. 1992, 2000; Mihailoff 1993). Các tổn thương ở cuống tiểu não giữa làm mất khả năng học tập với các CS thính giác, thị giác hoặc xúc giác (Solomon và cộng sự 1986; Lewis và cộng sự 1987). Tính hiệu quả của việc truyền dẫn sợi rêu như một CS đã được chứng minh bằng một loạt nghiên cứu sử dụng kích thích cầu não hoặc cuống tiểu não giữa khi một CS kết hợp với US ngoại vi (Steinmetz và cộng sự 1986; Rosen và cộng sự 1989; Steinmetz 1990; Tracy và cộng sự 1998, Hesslow và cộng sự 1999, Bao và cộng sự 2000, Freeman và Rabinak 2004, Freeman và cộng sự 2005b). Kích thích sợi rêu là giải pháp CS điều kiện hóa chớp mắt cực kỳ hiệu quả. Hơn nữa, các thông số hỗ trợ điều kiện hóa và giải trừ với CS sợi ngoại vi và sợi rêu rất giống nhau. Tất nhiên, kích thích điện có thể gây ra sự kích hoạt antidromic [ngược hướng, đối nghĩa với orthodromic] và kích hoạt (hoặc ức chế) các mục tiêu ngoài ý muốn. Mối lo ngại chung này đã được xoa dịu nhờ các thí nghiệm cho thấy sự loại bỏ các CR được hình thành do sự kích thích của sợi rêu từ các tổn thương hoặc sự bất hoạt của nhân xen trước (Steinmetz và cộng sự 1986; Freeman và Rabinak 2004; Freeman và cộng sự 2005b). Hesslow và các đồng nghiệp cũng đã thiết lập khả năng điều kiện hóa chớp mắt bằng CS kích thích sợi rêu kết hợp với chất chặn kênh natri trong cuống tiểu não giữa nằm xa điện cực kích thích để ngăn chặn sự kích hoạt antidromic của các neuron thân não (Hesslow và cộng sự 1999). Nói chung, các nghiên cứu về tổn thương và kích thích chỉ ra rằng thông tin CS, bất kể phương thức cảm giác nào, đều được dẫn tới tiểu não thông qua các sợi rêu.
Con đường thính giác CS
Các truyền dẫn vỏ thính giác tới nhân cầu não tương đối nổi bật, nhưng các tổn thương của vỏ não bao gồm cả vỏ thính giác không ngăn cản việc thu nhận hoặc duy trì khả năng điều kiện hóa chớp mắt trễ (Oakley và Russell 1972, 1977). Con đường CS thính giác dưới vỏ não ban đầu được Steinmetz và các đồng nghiệp nghiên cứu bằng cách sử dụng thỏ (Steinmetz và cộng sự 1987). Các nhân bên của cầu não được xác định là nguồn cung cấp sợi thính giác dạng rêu đến tiểu não bằng cách sử dụng các tổn thương sau luyện tập. Những con thỏ bị tổn thương điện phân ở nhân cầu não bên sau khi có được khả năng điều kiện hóa thính giác trong chớp mắt đã bị suy giảm nghiêm trọng. Tuy nhiên, những con thỏ này không bị suy yếu khi được thử nghiệm bằng CS thị giác. Một truyền dẫn từ nhân ốc tai phía bụng tới các cầu não bên đã được xác định và được cho là nguồn có khả năng cung cấp đầu vào thính giác có độ trễ ngắn tới các cầu não, điều này được nhìn thấy bằng các bản ghi điện trường do thính giác gợi lên. Dựa trên tập hợp phát hiện này, Steinmetz et al. (1987) kết luận rằng con đường thính giác CS có khả năng xảy ra nhất là sự truyền dẫn nhân ốc tai tới nhân cầu não bên nhưng thừa nhận rằng các đầu vào thính giác khác có thể đóng một vai trò nào đó.
Các nghiên cứu sau đó cho thấy rằng củ não dưới/colliculus và đồi thị thính giác cũng đóng một vai trò trong việc điều kiện hóa thính giác chớp mắt ở chuột. Các tổn thương một bên của củ dưới hoặc thể gối trong và các nhân đồi thị lân cận đối bên với mắt điều kiện hóa làm suy giảm nghiêm trọng khả năng điều kiện hóa chớp mắt trễ với trương lực CS (Halverson và Freeman 2006; Freeman và cộng sự 2007). Các tổn thương giới hạn ở phần lưng và phần bụng của thể gối trong không có tác dụng, nhưng các tổn thương của nhân đồi thị thính giác trong (medial auditory thalamic nuclei MATN), bao gồm phần trong của thể gối trong, nhân nội lá tủy sau và nhân trên thể gối, ngăn chặn hoàn toàn việc tiếp nhận điều kiện hóa thính giác chớp mắt. Những con chuột bị tổn thương MATN hoàn toàn không bị suy giảm khi được huấn luyện bằng CS thị giác, cho thấy rằng sự suy giảm đó là đặc trưng cho một phương thức. Các thí nghiệm theo dõi đường ngược và xuôi dòng cho thấy một truyền dẫn cùng bên đơn synap từ MATN đến nhân cầu não, và sự kích thích trong truyền dẫn này dưới dạng CS là đủ để tạo ra sự tiếp nhận nhanh chóng khả năng điều kiện hóa chớp mắt (Campolattaro và cộng sự 2007). Một nghiên cứu tiếp theo cho thấy nhân cầu não bên đối diện với mắt điều kiện hóa rất quan trọng trong việc chuyển tiếp thông tin thính giác CS từ MATN đến tiểu não (Halverson và Freeman 2010a). Do đó, con đường thính giác CS bao gồm các truyền song song từ nhân thân não thính giác phía dưới đến MATN đối diện với mắt điều kiện hóa, truyền dẫn cùng bên của chúng tới nhân cầu não bên và truyền dẫn sợi rêu đến vỏ tiểu não và nhân xen trước cùng bên với mắt điều kiện hóa ( Hình 2).
Các bản ghi thần kinh từ nhân cầu não trong quá trình điều kiện hóa thính giác cho thấy rằng các neuron cầu não chủ yếu biểu hiện các phản ứng có độ trễ ngắn với CS khi bắt đầu huấn luyện nhưng sau đó cho thấy sự gia tăng hoạt động vào cuối giai đoạn CS-US (Clark và cộng sự 1997; Bao và cộng sự 2000, Freeman và Muckler 2003, Campolattaro và cộng sự 2011). Quy trình liên quan đến học tập này bị loại bỏ do tiểu não ngừng hoạt động, cho thấy rằng phản hồi từ tiểu não thúc đẩy hoạt động của cầu não liên quan đến học tập (Cartford và cộng sự 1997; Clark và cộng sự 1997; Bao và cộng sự 2000). Một mô hình kết quả tương tự đã thu được với các bản ghi neuron thần kinh trong MATN đối bên với mắt điều kiện hóa (Halverson và cộng sự 2010). Các tế bào thần kinh MATN cũng thể hiện hoạt động liên quan đến học tập trong quá trình điều hòa (O’Connor và cộng sự 1997; Halverson và cộng sự 2010), và các bản ghi đồng thời của MATN và tế bào thần kinh khu vực phía trước đã chứng minh rằng sự gia tăng hoạt động liên quan đến học tập trong MATN theo sau sự gia tăng liên quan đến hoạt động của tiểu não trong các thử nghiệm huấn luyện, cho thấy rằng phản hồi của tiểu não có thể thúc đẩy hoạt động liên quan đến học tập ở đồi thị (Halverson và cộng sự 2010). Những phát hiện của các nghiên cứu sinh lý thần kinh này cho thấy rằng có một vòng phản hồi dương tính giữa tiểu não và đường dẫn truyền thần kinh thính giác của nó. Phản hồi dương tính này có thể tạo điều kiện thuận lợi cho tốc độ và tiệm cận của điều kiện hóa chớp mắt bằng cách tăng cường đầu vào CS tới tiểu não (Hình 3).
Con đường CS thị giác
Giống như con đường thính giác CS, sự tách biệt hoặc tổn thương của vỏ não cảm giác không ngăn cản việc thu nhận hoặc duy trì khả năng điều kiện hóa chớp mắt thị giác chậm (Hilgard và Marquis 1935, 1936; Oakley và Russell 1975, 1976, 1977).
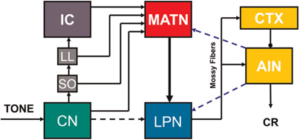 Hình 2. Con đường CS thính giác được giả thuyết để điều kiện hóa chớp mắt. Có các đầu vào song song vào nhân đồi thị thính giác trong (MATN) từ nhân ốc tai (CN), olive trên (SO), nhân của cảm giác bên (LL) và củ dưới (IC). Sau đó, đầu vào thính giác được dẫn từ MATN tới nhân cầu não bên (LPN), nhân này truyền tới vỏ tiểu não (CTX) và nhân xen trước (AIN). Các truyền dẫn phản hồi giả thuyết từ AIN đến LPN và MATN cũng được hiển thị (các đường đứt nét màu xanh lam).
Hình 2. Con đường CS thính giác được giả thuyết để điều kiện hóa chớp mắt. Có các đầu vào song song vào nhân đồi thị thính giác trong (MATN) từ nhân ốc tai (CN), olive trên (SO), nhân của cảm giác bên (LL) và củ dưới (IC). Sau đó, đầu vào thính giác được dẫn từ MATN tới nhân cầu não bên (LPN), nhân này truyền tới vỏ tiểu não (CTX) và nhân xen trước (AIN). Các truyền dẫn phản hồi giả thuyết từ AIN đến LPN và MATN cũng được hiển thị (các đường đứt nét màu xanh lam).
Hình 3. Vai trò giả thuyết được đưa ra của độ khả biến của đồi thị thính giác trong quá trình học tập của tiểu não. Các đầu vào kích thích được ghép nối từ nhân cầu não bên (LPN) và nhân olive dưới ở mặt sau (DAO) hội tụ trên các tế bào Purkinje ở vỏ tiểu não trong giai đoạn đầu của quá trình điều điều kiện hóa chớp mắt (trái). Quá trình học tập được bắt đầu bằng các sợi song song do CS kích hoạt đến các tế bào Purkinje gần như đồng thời với đầu vào sợi leo (cửa sổ). Không có độ khả biến liên quan đến học tập được thể hiện rõ ràng ở nhân xen (IPN) hoặc nhân đồi thị thính giác trung gian (MATN) trong các thử nghiệm huấn luyện ban đầu. 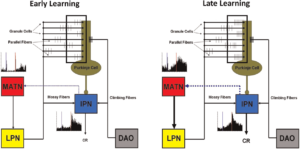 Các đầu vào kích thích được ghép nối từ LPN và DAO tiếp tục hội tụ trên các tế bào Purkinje sau khi quá trình học bắt đầu xuất hiện (phải). Quá trình học tập được tăng cường nhờ sự gia tăng (so với đào tạo ban đầu) hoạt động song song của sợi xảy ra gần như đồng thời với đầu vào sợi leo (cửa sổ) và hoạt động của sợi rêu trong IPN. Tính linh hoạt liên quan đến học tập rõ ràng được thể hiện rõ ràng trong IPN và MATN trong giai đoạn học tập sau này. Ngược lại, sự gia tăng phản hồi tiểu não kích thích liên quan đến học tập đối với MATN (đường chấm màu xanh) có thể làm tăng đầu ra MATN đến LPN và dẫn truyền sợi rêu tương ứng vào tiểu não để tạo điều kiện thuận lợi hơn cho việc học.
Các đầu vào kích thích được ghép nối từ LPN và DAO tiếp tục hội tụ trên các tế bào Purkinje sau khi quá trình học bắt đầu xuất hiện (phải). Quá trình học tập được tăng cường nhờ sự gia tăng (so với đào tạo ban đầu) hoạt động song song của sợi xảy ra gần như đồng thời với đầu vào sợi leo (cửa sổ) và hoạt động của sợi rêu trong IPN. Tính linh hoạt liên quan đến học tập rõ ràng được thể hiện rõ ràng trong IPN và MATN trong giai đoạn học tập sau này. Ngược lại, sự gia tăng phản hồi tiểu não kích thích liên quan đến học tập đối với MATN (đường chấm màu xanh) có thể làm tăng đầu ra MATN đến LPN và dẫn truyền sợi rêu tương ứng vào tiểu não để tạo điều kiện thuận lợi hơn cho việc học.
Phân tích sau đó của các đầu vào vỏ não và dưới vỏ não đối với nhân cầu não ở thỏ cho thấy một con đường CS thị giác song song bao gồm nhân gối bên (lateral geniculate nucleus LGN), lồi củ trên (superior colliculus SC), vỏ não thị giác (visual cortex VCTX) và nhân trước mái (Koutalidis et al. 1988). Chỉ riêng các tổn thương hai bên của LGN, SC, VCTX hoặc nhân trước mái/pretectal nuclei đã tạo ra suy giảm một phần; tổn thương của các cặp cấu trúc này gây ra tình trạng suy giảm nghiêm trọng hơn, nhưng chỉ những tổn thương kết hợp của tất cả các khu vực này mới ngăn chặn hoàn toàn việc điều kiện hóa chớp mắt bằng CS ánh sáng (Koutalidis et al. 1988). Sự kích thích của VCTX, SC và LGN ở bụng đối diện sẽ tạo phản xạ có điều kiện chớp mắt ở chuột và mỗi cấu trúc này có một truyền dẫn một chiều trực tiếp tới PN, có thể quan trọng đối với việc điều kiện hóa chớp mắt (Glickstein và cộng sự 1972; Graybiel 1974 Mower và cộng sự 1980, Legg và cộng sự 1989, Wells và cộng sự 1989, Halverson và cộng sự 2009). Các nhân trước mái cũng có truyền dẫn trực tiếp tới cả PN trong và ngoài (Weber và Harting 1980; Wells và cộng sự 1989), nhưng việc kích thích nhân trước mái trước không phải là một CS hiệu quả để điều kiện hóa chớp mắt (Campolattaro và cộng sự 2007). Một nghiên cứu gần đây đã cung cấp sự hỗ trợ bổ sung cho mô hình song song của con đường CS thị giác đồng thời gợi ý rằng chỉ một tập hợp con các đầu vào thị giác tới cầu não là cần thiết cho việc điều kiện hóa chớp mắt (Haverson và Freeman 2010b). Halverson và Freeman (2010b) phát hiện ra rằng sự bất hoạt đơn phương của nhân cầu não trong đối diện với mắt điều kiện hóa đã loại bỏ khả năng duy trì điều kiện hóa chớp mắt thị giác. Vùng cầu não trong bị bất hoạt sẽ nhận đầu vào đơn synap từ LGN bụng cùng bên và nhân của bó thị giác (Haverson và Freeman 2010b). Kết quả của nghiên cứu Halverson và Freeman (2010b) cho thấy rằng con đường CS thị giác bao gồm đầu vào song song đơn phương từ LGN bụng và nhân của bó thị giác đến các cầu não trong và dẫn truyền sợi rêu của nó tới vỏ tiểu não và nhân xen trước (Hình 4).
Hình 4. Con đường giả thuyết CS thị giác cần thiết cho việc duy trì điều kiện hóa chớp mắt. Các đầu vào từ võng mạc đến nhân gối bụng bên (ventral lateral geniculate LGNv) và nhân của đường thị giác (optic tract OT) được chuyển tiếp song song với vỏ tiểu não (CTX) và nhân xen trước (AIN) thông qua nhân cầu não trong (MPN). 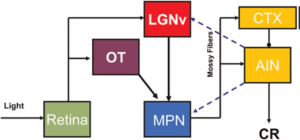 Các truyền dẫn phản hồi giả thuyết từ AIN đến MPN và LGNv cũng được hiển thị (các đường đứt nét màu xanh lam).
Các truyền dẫn phản hồi giả thuyết từ AIN đến MPN và LGNv cũng được hiển thị (các đường đứt nét màu xanh lam).
Con đường CS cảm giác thân thể
Kích thích rung kích hoạt các đường hướng tâm đến tủy sống, hướng tới các nhân cột sau. Các nhân của tủy sống sau đó chiếu tới nhân cầu não trong (Kosinski và cộng sự 1986a,b, 1988). Truyền dẫn của nhân nêm (chi trước và đầu thân) đối xứng với truyền dẫn của nhân nêm (chi sau và đuôi thân). Các nghiên cứu trước đây trên thỏ và chồn sương đã chỉ ra rằng việc điều kiện hóa chớp mắt bằng cách kích thích cơ thể với rung động hoặc sốc điện yếu phụ thuộc vào cuống tiểu não giữa (Solomon và cộng sự 1986; Lewis và cộng sự 1987; Hesslow và cộng sự 1999). Do đó, mặc dù có những dẫn truyền trực tiếp từ nhân cột sau đến tiểu não thông qua cuống tiểu não dưới (Bengtsson và Jorntell 2009), con đường CS có khả năng nhất đối với rung CS là dẫn truyền nhân cột sau tới nhân cầu não trong.
Tóm tắt mạch đầu vào kích thích có điều kiện
Các đường dẫn đầu vào kích thích có điều kiện cần thiết cho việc điều kiện hóa chớp mắt trễ chưa được nghiên cứu chi tiết, nhưng dữ liệu mới chỉ ra rằng các đầu vào từ nhiều nguồn dưới vỏ não khác nhau được gửi đến nhân nền cầu não và sau đó qua cuống tiểu não giữa đến vỏ tiểu não và nhân sâu. Các phát hiện cho đến nay cho thấy rằng không có cấu trúc mạch chung trong các con đường CS: Con đường thính giác đến cầu não chủ yếu là nối tiếp trong khi con đường thị giác là song song. Một điểm khác biệt nữa về mạch thần kinh là các con đường thính giác và thị giác bao gồm các đầu vào đồi thị tới cầu não, trong khi con đường cảm giác thân thể phát sinh từ nhân tủy. Cũng có thể có sự khác biệt giữa các con đường CS về lượng phản hồi tiểu não và tính khả biến liên quan đến học tập.
Mô hình vi tính toán điều kiện hóa chớp mắt
Tiến bộ đáng kể đã được thực hiện trong việc xác định vị trí lưu trữ bộ nhớ và độ khả biến của synap cũng như mạch thần kinh đầu vào và đầu ra cần thiết và đủ để duy trì điều kiện hóa chớp mắt (Hình 5). Tuy nhiên, các tương tác động phức tạp giữa các phần tử của mạch thần kinh không thể được mô tả trong các mô hình tĩnh như mô hình được trình bày trong Hình 5. Các mô hình vi tính toán rất hữu ích trong việc mô tả bản chất động của việc học tập ở tiểu não và để tạo ra các giả thuyết có thể kiểm chứng được cho nghiên cứu thực nghiệm (Moore et cộng sự 1986, 1989; Desmond và Moore 1988; Berthier và cộng sự 1991; Bartha và Thompson 1992; Fiala và cộng sự 1996; Mauk và Donegan 1997; Kenyon và cộng sự 1998a,b; Medina và Mauk 1999, 2000; Medina và cộng sự 2000, 2001, Ohyama và cộng sự 2003, Mauk và Buonomano 2004, Lepora và cộng sự 2010). Một phương pháp vi tính toán đặc biệt hứa hẹn là mô phỏng tiểu não trên máy tính của Mauk và các đồng nghiệp. Mô phỏng này bao gồm tất cả các loại neuron trong tiểu não, theo tỷ lệ chính xác, với các mối quan hệ kết nối synap tương ứng. Mô phỏng bao gồm các tương tác bổ sung giữa các neuron trong các thử nghiệm huấn luyện cũng như các thay đổi theo giai đoạn của hoạt động trong quá trình trình bày các kích thích. Cơ chế khả biến bao gồm những thay đổi hai chiều trong các synao sợi song song với tế bào Purkinje và các synap sợi rêu với các neuron xen trước. Mô hình mô phỏng chính xác nhiều phát hiện được mô tả ở trên nhưng cũng đã tạo ra một số giả thuyết mới đã được nghiên cứu thực nghiệm. Ví dụ, mô phỏng bao gồm phản hồi ức chế trương lực và pha từ tiểu não đến olive dưới. Dự đoán của mô hình là mức độ kích hoạt trong các sợi leo đóng vai trò quan trọng trong việc xác định liệu các sợi thần kinh song song có tạo synap với các tế bào Purkinje trải qua LTP, LTD hay duy trì trạng thái hiện tại của chúng hay không. Theo đó, sự giải trừ được thiết lập bằng cách giảm hoạt động của sợi leo xuống dưới mức “cân bằng” tạo ra LTP, một dự đoán duy nhất của mô phỏng đã được xác minh bằng cách ngăn chặn phản hồi ức chế đối với olive dưới trong quá trình huấn luyện giải trừ (Medina và cộng sự 2002). Ví dụ này minh họa cách một mô hình thời gian thực bao gồm các tương tác động đã biết giữa các phần tử của mạch có thể mô phỏng chính xác các cơ chế học tập và tạo ra các giả thuyết. Việc phát triển hơn nữa các mô phỏng máy tính bao gồm các tương tác với mạch đầu vào CS và các ảnh hưởng điều chỉnh não trước có thể tạo điều kiện hiểu biết đầy đủ hơn về các cơ chế duy trì điều kiện hóa cơ bản.
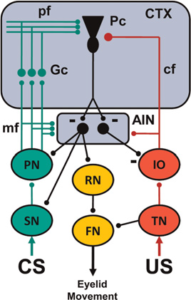 Hình 5. Lược đồ đơn giản hóa của mạch thần kinh làm cơ sở cho việc điều kiện hóa chớp mắt. Nhân xen trước tiểu não (AIN) và tế bào Purkinje (Pc) trong vỏ tiểu não (CTX) nhận đầu vào hội tụ từ các đường thần kinh kích thích có điều kiện (CS, xanh lá cây) và kích thích không điều kiện (US, đỏ). Con đường cho các CS khác nhau bao gồm nhân cảm giác dưới vỏ não (SN), nhân cầu não nền (PN), chiếu sợi rêu (mf) tới AIN và tế bào hạt vỏ não (Gc) và dẫn truyền sợi song song (pf) đến tế bào Purkinje. Con đường US bao gồm nhân ba hạt (TN), phần phụ ở mặt lưng của olive dưới (IO) và phần nhô ra của sợi leo (cf) tới AIN và Pc. Con đường đầu ra để thực hiện phản xạ có điều kiện (màu cam) bao gồm truyền dẫn AIN tới nhân đỏ (RN) và truyền dẫn của nó tới nhân vận động mặt (FN) gây ra việc đóng mí mắt. Phản xạ không điều kiện được tạo ra bằng cách kích hoạt TN, sau đó kích hoạt FN. Các dẫn truyền phản hồi từ AIN đến PN, SN và IO điều chỉnh đầu vào CS và US để tạo điều kiện thuận lợi cho việc tiếp nhận và duy trì độ khả biến trong tiểu não. Các synap ức chế được mô tả bằng dấu trừ. Tất cả các synap khác đều là kích thích.
Hình 5. Lược đồ đơn giản hóa của mạch thần kinh làm cơ sở cho việc điều kiện hóa chớp mắt. Nhân xen trước tiểu não (AIN) và tế bào Purkinje (Pc) trong vỏ tiểu não (CTX) nhận đầu vào hội tụ từ các đường thần kinh kích thích có điều kiện (CS, xanh lá cây) và kích thích không điều kiện (US, đỏ). Con đường cho các CS khác nhau bao gồm nhân cảm giác dưới vỏ não (SN), nhân cầu não nền (PN), chiếu sợi rêu (mf) tới AIN và tế bào hạt vỏ não (Gc) và dẫn truyền sợi song song (pf) đến tế bào Purkinje. Con đường US bao gồm nhân ba hạt (TN), phần phụ ở mặt lưng của olive dưới (IO) và phần nhô ra của sợi leo (cf) tới AIN và Pc. Con đường đầu ra để thực hiện phản xạ có điều kiện (màu cam) bao gồm truyền dẫn AIN tới nhân đỏ (RN) và truyền dẫn của nó tới nhân vận động mặt (FN) gây ra việc đóng mí mắt. Phản xạ không điều kiện được tạo ra bằng cách kích hoạt TN, sau đó kích hoạt FN. Các dẫn truyền phản hồi từ AIN đến PN, SN và IO điều chỉnh đầu vào CS và US để tạo điều kiện thuận lợi cho việc tiếp nhận và duy trì độ khả biến trong tiểu não. Các synap ức chế được mô tả bằng dấu trừ. Tất cả các synap khác đều là kích thích.
Điều kiện duy trì điều kiện hóa
Một số hệ thống thần kinh ảnh hưởng đến khả năng duy trì điều kiện hóa nhưng không cần thiết cho việc tiếp nhận hoặc lưu giữ. Một cuộc thảo luận đầy đủ về vấn đề này nằm ngoài phạm vi của bài đánh giá này, nhưng điều quan trọng cần lưu ý là vùng hippocampal theta, amygdala, vỏ não cảm giác và norepinephrine đầu vào cho mạch học tập được mô tả ở trên điều chỉnh việc tiếp nhận điều kiện hóa chớp mắt (Berry và Thompson 1979; Neufeld và Mintz 2001, Allen và cộng sự 2002, Case và cộng sự 2002, Cartford và cộng sự 2004, Lee và Kim 2004, Blankenship và cộng sự 2005, Taub và Mintz 2010). Một mô hình thực sự toàn diện về điều kiện hóa chớp mắt sẽ kết hợp các cơ chế điều biến bên cạnh mạch thiết yếu.
Hướng đi tương lai trong nghiên cứu điều kiện hóa chớp mắt
Mặc dù các cơ chế thần kinh làm duy trì quá trình điều kiện hóa chớp mắt đã được mô tả đầy đủ hơn bất kỳ hình thức học tập nào khác của động vật có vú, nhưng có một số vấn đề quan trọng cần được nghiên cứu thêm. Ở cấp độ mạch, các đường dẫn CS chưa được xác định đầy đủ cho bất kỳ phương thức cảm giác nào cũng như không có bản chất tương tác giữa một số đầu vào CS đã biết và đầu ra tiểu não (Hình 3). Một vấn đề cấp độ mạch quan trọng khác là mô tả đặc điểm của các cơ chế thần kinh làm cơ sở điều chế não trước duy trì điều kiện hóa của amygdala, hồi hải mã và vỏ não (Berry và Thompson 1979; Neufeld và Mintz 2001; Allen và cộng sự 2002; Case và cộng sự 2002; Cartford và cộng sự 2004; Lee và Kim 2004; Blankenship và cộng sự 2005; Taub và Mintz 2010). Có bằng chứng cho thấy các hệ thống này tạo điều kiện thuận lợi cho việc duy trì điều kiện hóa chớp mắt, nhưng chính xác làm thế nào đạt được sự hỗ trợ này hiện vẫn còn là bí ẩn.
Các cơ chế tế bào và phân tử làm cơ sở cho việc học tập của não cũng cần được kiểm tra thêm. Các cơ chế khả biến đã được đưa ra giả thuyết và những thay đổi synap trong tiểu não đã được xác định, nhưng các cơ chế tế bào và phân tử làm cơ sở cho những thay đổi liên quan đến học tập trong các synap vẫn chưa được biết. Các cơ chế cảm ứng độ khả biến của synap trong tiểu não đã được mô hình hóa trong các chế phẩm khử nhưng chưa được xác minh in vivo trong quá trình điều kiện hóa. Ví dụ, mô phỏng tiểu não của Mauk và các đồng nghiệp dự đoán rằng việc tạo ra LTD ở vỏ não là được phép và do đó có trước việc tạo ra LTP ở nhân xen trước (Mauk và Donegan 1997; Pugh và Raman 2008, 2009), nhưng đồng thời các bản ghi cho thấy rằng việc tạm dừng trong các nhọn sóng đơn giản của tế bào Purkinje dẫn đến sự phát triển của hoạt động xen tăng lên trong quá trình tiếp nhận ban đầu chưa được tiến hành. Hơn nữa, mối quan hệ cơ học giữa những thay đổi về số lượng hoặc hình thái synap và những thay đổi về hiệu quả của con đường vẫn chưa được xác định. Cuối cùng, vai trò của các neuron trung gian vỏ tiểu não trong điều kiện hóa chớp mắt chưa được xem xét chi tiết nhưng giờ đây khả thi hơn khi cấu hình kích hoạt in vivo của các neuron trung gian được xác định đã được mô tả (Rigrok và cộng sự 2011). Các vấn đề rộng lớn chưa được giải quyết được mô tả trong phần này chỉ ra rằng nghiên cứu về duy trì điều kiện hóa chớp mắt sẽ tiếp tục là một lĩnh vực nghiên cứu thú vị và tích cực trong tương lai gần.



Đăng nhập để bình luận.