
Các dạng chớp mắt khác nhau và cách kiểm soát hai giai đoạn của chúng



- Bs Đỗ Thị Thúy Anh
Tóm tắt. Mục đích của bài viết này là kiểm tra chuyển động học (kinematic) của chớp mắt và cơ sở thần kinh của chớp mắt được gợi lên theo phản xạ bởi các loại kích thích khác nhau. Chuyển động học của chuyển động của mí mắt trên và phản ứng điện cơ của cơ nâng mí mắt và cơ vòng mắt đã được ghi lại ở thỏ trong các chớp mắt sinh ba và gợi lên bằng thị giác. Chúng tôi thấy rằng có sự khác biệt cơ bản về mặt chuyển động học giữa các lần chớp mắt. Nháy mắt để phản ứng với một luồng khí được thực hiện nhanh hơn và đạt được vận tốc cao hơn so với chớp mắt có biên độ bằng nhau để phản ứng với một tia sáng. Hai dạng chớp mắt phản xạ là do sự khác biệt về bản chất và thời gian hoạt động của các neuron vận động của cơ mí mắt (lid muscle) đối kháng. Tuy nhiên, hầu hết các đặc điểm của việc điều khiển thần kinh chớp mắt đều giống nhau ở cả hai kiểu chớp mắt phản xạ. Quan trọng nhất, có vẻ như chớp mắt được tạo ra bởi sự điều khiển thần kinh hai giai đoạn, một thành phần sớm được lập trình sẵn và một thành phần muộn được kiểm soát kích thích.
Giới thiệu
Nháy mắt được công nhận là một xét nghiệm thần kinh quan trọng về tính toàn vẹn của thân não. Nhiều nghiên cứu gần đây đã xem xét các khía cạnh liên quan của chớp mắt: chuyển động của mắt khi chớp mắt (Collewijn và cộng sự 1985; Evinger et a1. 1984), thị lực khi chớp mắt (Manning et al. 1983; Riggs et al. 1981), và chảy nước mắt (Doane 1980). Nháy mắt đã đóng vai trò như một hành vi mẫu cho các hình thức học tập đơn giản: học tập kết hợp/associative learning [phản xạ có điều kiện] (McCormick và Thompson 1984; Yeo và cộng sự 1985), điều chỉnh phản xạ/reflex modification (Hoffman và Ison 1980; Uhlrich và cộng sự 1983), và tính linh hoạt thích ứng/adaptive plasticity (Evinger và Manning 1985). Bất chấp sự quan tâm gần đây đến chớp mắt, một số đặc tính cơ bản của chúng vẫn chưa được hiểu rõ.
Một giả định cơ bản của hầu hết các nghiên cứu là chớp mắt là một chuyển động thống nhất và một khi được gợi ra thì tất cả các chớp mắt đều giống nhau. Tuy nhiên, nhiều con đường hướng tâm có thể gợi lên một chớp mắt và độ trễ chớp mắt phụ thuộc vào bản chất của kích thích khơi gợi (Hopf và cộng sự 1973; Kugelberg 1952; Rushworth 1962; Yates và Brown 1981; Zametkin và cộng sự 1979). Các nghiên cứu trước đây không kiểm tra kỹ lưỡng chuyển động học của chớp mắt hoặc cơ sở thần kinh của chớp mắt được gợi lên bởi các kích thích khác nhau, mặc dù Doane (1980) đã lưu ý một số khác biệt về vận tốc mi mắt giữa chớp mắt tình nguyện và chớp mắt tự phát (spontaneous blinks) ở các đối tượng là con người.
Mặc dù tất cả các lần chớp mắt đều phụ thuộc vào cùng một cơ mí mắt, nhưng chuyển động của mí mắt không nhất thiết phải giống hệt nhau. Ví dụ, các dạng chuyển động cơ bản khác nhau rõ rệt (ví dụ: điều chỉnh tư thế) xảy ra do sự thay đổi về thời gian co bóp trong một tập hợp cơ (để xem lại, hãy xem Nashner và McCollum 1985). Nghiên cứu hiện tại chứng minh rằng chớp mắt do dây sinh ba và chớp mắt được gợi lên bằng thị giác ở thỏ khác biệt về mặt chuyển động học và điều này là kết quả của sự khác biệt về bản chất và thời gian co cơ. Hơn nữa, dữ liệu của chúng tôi cho thấy rằng việc coi một chớp mắt của một cá nhân như một chuyển động thống nhất sẽ che khuất một khía cạnh quan trọng về cách hệ thống thần kinh tổ chức các chớp mắt và chuyển động nói chung. Ví dụ, các chuyển động đơn giản khác như chuyển động mắt theo đuổi trơn tru thực sự bao gồm hai thành phần, thành phần có độ trễ ngắn để bắt đầu chuyển động và thành phần có độ trễ dài hơn giúp điều chỉnh phản ứng với các đặc điểm của kích thích (Lisberger và Westbrook 1985). Nghiên cứu hiện tại gợi ý rằng việc điều khiển thần kinh của phản xạ chớp mắt hoạt động theo kiểu hai phần tương tự. Trong cả hai loại chớp mắt phản xạ, chuyển động chớp mắt ban đầu được lập trình trước trong khi thành phần sau của chớp mắt được kiểm soát kích thích.
Vật liệu và phương pháp
Thủ tục phẫu thuật
Đối tượng của thí nghiệm này là sáu con thỏ bạch tạng New Zealand trưởng thành (2 đến 3,5 kg). Dưới gây mê toàn thân (ketamine, acepromazine, xylazine; xem Shaw và Alley, 1982) và trong điều kiện vô trùng, các cặp điện cực được nhúng vào cơ mí mắt của bốn con thỏ để làm điện cực ghi điện cơ (EMG). Các cơ chính liên quan đến chớp mắt là cơ cơ vòng mí mắt/ orbicularis oculi (OO), một cơ vòng bao quanh mắt và đóng mí mắt, và cơ nâng mi mắt/ levator palpebrae (LP), nằm phía trên cơ thẳng trên và bám tận vào mí mắt trên, nâng cao mi mắt trên. Các điện cực EMG được đặt chính giữa vào OO của mí mắt trên, cách mép mí mắt vài mm và vào LP, gần bụng cơ. Đầu mỗi dây thép không gỉ sợi đơn, phủ Teflon (trần 0,003 inch; bọc 0,0045 inch) được vạch ra 1 mm, luồn vào kim tiêm dưới da cỡ 27, móc ở đầu kim rồi trượt kim vào cơ và rút ra, để lại dây điện cực được cấy ghép. Một sợi dây bạc nằm dọc theo hộp sọ được dùng làm điện cực nối đất. Vòng quấn dây thần kinh kích thích mãn tính chứa hai dây loại EMG được chế tạo từ ống Teflon (ID = 0,034“, OD = 0,04“) và được cấy vào hai con thỏ xung quanh nhánh trên ổ mắt của dây thần kinh sinh ba, trước đó đã được loại bỏ khỏi mô xung quanh . Các dây EMG, dây nối đất và dây quấn dây thần kinh đi qua da về phía đỉnh đầu và được kết nối với các phích cắm trên vương miện acrylic nha khoa được cố định vào hộp sọ bằng sáu ốc vít. Vương miện được gắn vào một thanh cố định trong thiết bị ghi để giữ đầu trong quá trình thu thập dữ liệu. Thỏ được dùng thuốc giảm đau sau phẫu thuật để giảm thiểu sự khó chịu. Thỏ tham gia thí nghiệm lần đầu tiên khoảng 1 tuần sau khi phẫu thuật, sau khi hồi phục hoàn toàn. Không có dấu hiệu nào cho thấy các quy trình này ảnh hưởng đến việc chớp mắt bình thường dưới bất kỳ hình thức nào.
Vị trí mí mắt
Trong quá trình thí nghiệm, một monitor nhạy cảm với vị trí mí mắt được dán vào mí mắt trên để đo vị trí của mí mắt trong mặt phẳng thẳng đứng. Một đầu của một đòn bẩy đối trọng, nhẹ được gắn vào một sợi tơ được cố định vào nắp gần giữa mép mí mắt trên (Evinger et al. 1984). Khi mí nâng lên và hạ xuống, một đi-ốt phát sáng ở đầu đối diện của cần di chuyển qua bộ dò vị trí cảm quang (United Detector Technology, SC-50). Đầu ra của bộ dò vị trí được khuếch đại khác nhau (Tektronix AM502; băng thông có điểm -3 dB tại DC và 1 KHz). Sự phân biệt tương tự (100 Hz, -3 dB) của đầu ra máy dò cung cấp thước đo vận tốc mí mắt. Hệ thống được hiệu chỉnh sau mỗi phiên bằng cách di chuyển cần gạt qua những khoảng cách đã biết. Vì thỏ có ít chuyển động mắt hoặc chớp mắt tự phát nên các bản ghi vị trí mí hoặc bản ghi điện cơ đều không bị nhiễu do chuyển động mắt không mong muốn.
Kích thích gợi lên chớp mắt
Các thí nghiệm được thiết kế để gợi ra những chớp mắt phản xạ thông qua con đường hướng tâm thị giác hoặc sinh ba. Tất cả các con thỏ đều chớp mắt trước những tia sáng rực rỡ từ đèn halogen (American Optical). Ánh sáng truyền qua một bó sợi quang và hướng vào mắt bằng một thấu kính để chiếu sáng một khu vực có đường kính khoảng 2,5 cm. Độ chói, được đo bằng quang kế Tektronix J16 với đầu dò J623, thay đổi từ 350 đến 21950 foot-Lambert. Một màn trập điện tử tác dụng nhanh (Uniblitz) xen kẽ vào đường dẫn ánh sáng có thời lượng đèn flash được kiểm soát từ 25 đến 500 mili giây. Thời gian kích thích ánh sáng đã được xác nhận bằng tế bào quang điện. Ngay cả sau hàng trăm thử nghiệm, việc tự kích hoạt màn trập cũng không gây ra một chớp mắt nào.
Tất cả các con thỏ cũng chớp mắt để phản ứng với những luồng không khí rời rạc hướng vào mắt và vùng quanh ổ mắt. Khí nén được dẫn qua ống cao su đến lỗ hẹp trong pipet nhựa (dài 5 mm, đường kính <1 mm ở đầu) cách mắt 30 mm. Áp suất không khí tại nguồn thay đổi từ 4 đến 17 lbs/sq in. Một van điện từ (Van chung) vận hành bằng điện tử có thời gian điều khiển luồng khí, dao động từ 25 đến 500 ms. Để theo dõi thời gian khởi phát và thời gian kích thích, một đoạn ống giống hệt ống dẫn đến mắt và chia sẻ cùng một nguồn áp suất được hướng về phía micrô cách xa 30 mm. Riêng âm thanh của tiếng thổi hơi không gây ra một chớp mắt. Với một ngoại lệ hiếm hoi, mỗi kích thích không khí và ánh sáng chỉ gây ra một chớp mắt duy nhất.
Cuối cùng, chớp mắt được tạo ra bởi các xung điện áp không đổi truyền qua dây quấn dây thần kinh trên ổ mắt ở thỏ được cấy ghép. Các xung nhấp nháy kéo dài 100 qs và điện áp dao động từ 20 đến 50 V (Bộ đếm thời gian WPI và bộ cách ly kích thích). Điện áp được tăng dần từ 0 V cho đến khi xảy ra chớp mắt. Sự kích thích không bao giờ vượt quá ngưỡng 1,5 lần và thỏ không bao giờ có dấu hiệu khó chịu.
Thu thập dữ liệu
Những con thỏ tỉnh táo, im lặng đã được thử nghiệm trong các phiên từ 1 đến 1 và 1/2 giờ trong đó thời gian kích thích hoặc biên độ kích thích khác nhau. Trong mỗi phiên, đầy đủ các kích thích được trình bày theo các khối được sắp xếp ngẫu nhiên từ 9 đến 12 thử nghiệm. Các khối thử nghiệm luồng khí airpuff xen kẽ với các khối thử nghiệm đèn flash. Máy tính kiểm soát thời gian giữa các lần thử nghiệm thay đổi ngẫu nhiên từ 12 đến 18 giây. Phản ứng chớp mắt cho thấy ít quen thuộc với khoảng thời gian giữa các lần thử nghiệm này. Việc lặp lại các điều kiện không khí và ánh sáng đầu tiên vào cuối phiên đã cung cấp ước tính về sự thay đổi của dữ liệu trong quá trình thử nghiệm. Có rất ít hoặc không có sự khác biệt giữa dữ liệu EMG hoặc dữ liệu mí mắt được thu thập khi bắt đầu và kết thúc thử nghiệm. Một số con thỏ đã được sử dụng nhiều lần trong cùng một thí nghiệm hoặc tham gia vào cả thí nghiệm về biên độ và thời gian. Thỏ 1 và 6 được thử nghiệm với phạm vi kích thích hạn chế hơn so với các thỏ khác.
Việc quan tâm đến sự thoải mái của thỏ là rất quan trọng vì sự khó chịu hoặc khó chịu sẽ dẫn đến những phản ứng không đáng tin cậy và khiến thỏ nhắm mắt lại, điều này sẽ kết thúc thí nghiệm.
Phân tích dữ liệu
Dữ liệu chuyển động của mí và EMG từ mỗi thử nghiệm được lưu trữ trên băng từ tương tự (máy ghi thiết bị Vetter 8 kênh) để phân tích ngoại tuyến sau này. Phản hồi EMG đã được lọc (băng thông có điểm -3 dB ở 100 Hz và 3 KHz). Dữ liệu được lưu trữ và phân tích trên PC IBM (chuyển đổi A/D với độ chính xác 12 bit, 1000 Hz/kênh cho vị trí và vận tốc và 6000 Hz/kênh cho EMG). Máy tính hiển thị từng bản ghi nháy mắt và chọn điểm bắt đầu, khoảng lệch tối đa và vận tốc tối đa của chuyển động hạ mí xuống, mặc dù người dùng có thể sửa đổi các giá trị này nếu cần. Đối với các thử nghiệm với bản ghi EMG, máy tính đã tính tích phân của hoạt động EMG đã được chỉnh sửa và lọc (i = 6,5 ms. Dữ liệu chớp mắt thu được được phân tích riêng lẻ hoặc lấy trung bình, bỏ qua chớp mắt đầu tiên trong mỗi thử nghiệm để loại bỏ bất kỳ hiệu ứng giật mình nào.
Kết quả
Sự khác biệt giữa chớp mắt do ánh sáng và chớp mắt do không khí
Nháy mắt của thỏ để phản ứng với kích thích sinh ba có độ trễ ngắn hơn so với chớp mắt khi phản ứng với kích thích thị giác (không khí, 25 đến 30 ms; ánh sáng, 80 đến 100 ms) như đã báo cáo trước đây đối với các loài khác (mèo: Hiraoka và Shimamura 1977; Lindquist và Martensson 1970, chuột: Hall và Hicks 1973, con người: Ongerboer de Visser 1980; Ongerboer de Visser và cộng sự 1977; Rushworth 1962; Shahani và Young 1973). Tuy nhiên, kiểm tra kỹ hơn cho thấy sự khác biệt về mặt chuyển động học chưa được báo cáo trước đây giữa các chớp mắt được tạo ra bởi các kích thích sinh ba và thị giác.
Trong quá trình hạ mi mắt với các chớp mắt có biên độ bằng nhau, mi mắt trên đạt được vận tốc và gia tốc lớn hơn đáng kể khi phản ứng với một luồng khí so với một nháy sáng nhẹ (Hình 1). 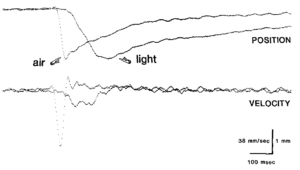 Dấu vết vận tốc chứng minh rằng chớp mắt do không khí tạo ra đạt được vận tốc mí hướng xuống lớn hơn đáng kể so với chớp mắt do ánh sáng tạo ra có biên độ bằng nhau. Sự khác biệt chuyển động học này kéo dài qua các chớp mắt ở mọi biên độ. Điều này được thấy rõ trong các biểu đồ về thời lượng chớp mắt, thời gian từ khi bắt đầu chuyển động của mí cho đến khi mí đạt được độ lệch hướng xuống tối đa và vận tốc mí tối đa, vận tốc cực đại đạt được trong quá trình đóng mí, như một hàm của biên độ chớp mắt, mức tối đa sự lệch của mí trong chớp mắt (Hình 2).
Dấu vết vận tốc chứng minh rằng chớp mắt do không khí tạo ra đạt được vận tốc mí hướng xuống lớn hơn đáng kể so với chớp mắt do ánh sáng tạo ra có biên độ bằng nhau. Sự khác biệt chuyển động học này kéo dài qua các chớp mắt ở mọi biên độ. Điều này được thấy rõ trong các biểu đồ về thời lượng chớp mắt, thời gian từ khi bắt đầu chuyển động của mí cho đến khi mí đạt được độ lệch hướng xuống tối đa và vận tốc mí tối đa, vận tốc cực đại đạt được trong quá trình đóng mí, như một hàm của biên độ chớp mắt, mức tối đa sự lệch của mí trong chớp mắt (Hình 2). 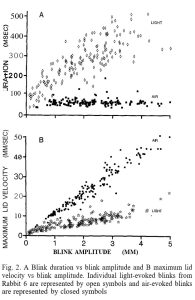 Các giá trị của độ dốc và điểm chặn y (y-intercept) để có các đường thẳng phù hợp nhất với dữ liệu khác nhau giữa các con thỏ, nhưng mẫu kết quả cơ bản đúng với tất cả các con thỏ (Bảng 1). Đối với mối quan hệ thời gian và biên độ, hệ số chặn y trung bình của ánh sáng lớn hơn khoảng 1,5 lần so với không khí và hệ số góc đối với ánh sáng xấp xỉ ba lần so với hệ số góc của không khí (trung bình chỉ lớn hơn 0). Các điểm chặn y không khác biệt đáng kể đối với mỗi con thỏ (ví dụ: Hình 2A), nhưng sự khác biệt về độ dốc được giữ ở mỗi con vật (so sánh độ lớn của độ lệch chuẩn độ dốc với độ lệch chuẩn chặn Y trong Bảng 1). Do đó, chớp mắt do ánh sáng kéo dài hơn ngay cả với những chớp mắt nhỏ và khi biên độ chớp mắt tăng lên, thời lượng của chớp mắt do ánh sáng tăng nhanh hơn thời lượng của chớp mắt do không khí. Thời lượng của các lần chớp mắt do ánh sáng tạo ra thay đổi nhiều hơn, nhưng hầu như luôn vượt quá thời lượng của các lần chớp mắt do không khí có biên độ bằng nhau.
Các giá trị của độ dốc và điểm chặn y (y-intercept) để có các đường thẳng phù hợp nhất với dữ liệu khác nhau giữa các con thỏ, nhưng mẫu kết quả cơ bản đúng với tất cả các con thỏ (Bảng 1). Đối với mối quan hệ thời gian và biên độ, hệ số chặn y trung bình của ánh sáng lớn hơn khoảng 1,5 lần so với không khí và hệ số góc đối với ánh sáng xấp xỉ ba lần so với hệ số góc của không khí (trung bình chỉ lớn hơn 0). Các điểm chặn y không khác biệt đáng kể đối với mỗi con thỏ (ví dụ: Hình 2A), nhưng sự khác biệt về độ dốc được giữ ở mỗi con vật (so sánh độ lớn của độ lệch chuẩn độ dốc với độ lệch chuẩn chặn Y trong Bảng 1). Do đó, chớp mắt do ánh sáng kéo dài hơn ngay cả với những chớp mắt nhỏ và khi biên độ chớp mắt tăng lên, thời lượng của chớp mắt do ánh sáng tăng nhanh hơn thời lượng của chớp mắt do không khí. Thời lượng của các lần chớp mắt do ánh sáng tạo ra thay đổi nhiều hơn, nhưng hầu như luôn vượt quá thời lượng của các lần chớp mắt do không khí có biên độ bằng nhau.
Đối với liên quan vận tốc và biên độ (Hình 2B), giao điểm y đối với cả ánh sáng và không khí là tương tự nhau, thường chỉ lớn hơn 0, nhưng hệ số góc đối với không khí lớn gần gấp đôi hệ số góc đối với ánh sáng. Do đó, trong cả hai điều kiện, vận tốc mí tăng tuyến tính theo biên độ chớp mắt, nhưng vận tốc mí tối đa tăng với tốc độ nhanh hơn đối với chớp mắt do không khí tạo ra. Do sự thay đổi về giá trị độ dốc và điểm chặn y giữa các thỏ, nên sự khác biệt giữa chức năng không khí và ánh sáng là nổi bật nhất khi so sánh với thỏ. Tuy nhiên, sau khi kết hợp dữ liệu từ tất cả các con thỏ (Bảng 1)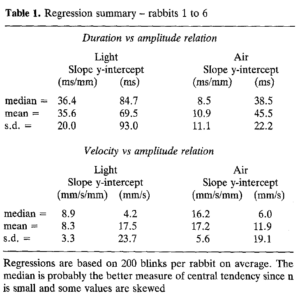 , sự khác biệt giữa ánh sáng và không khí vẫn rõ ràng.
, sự khác biệt giữa ánh sáng và không khí vẫn rõ ràng.
Sự khác biệt trong hoạt động cơ bắp
Chuyển động học khác nhau của chớp mắt do ánh sáng và không khí gây ra phải là do sự khác biệt trong hoạt động của cơ mí mắt. Ở mí mắt trên, sự co lại của cơ nâng mí mắt (LP) nâng mí mắt lên trong khi sự co lại của cơ vòng mi mắt (OO) cộng với lực căng thụ động hướng xuống làm mi mắt thấp xuống (xem Evinger et al. 1984 để đánh giá). Nháy mắt xảy ra khi các cơ hoạt động tương hỗ hoạt động phối hợp để LP thư giãn và OO co lại, tạo ra giai đoạn chớp mắt, sau đó hoạt động OO dừng lại và hoạt động LP tiếp tục, tạo ra giai đoạn chớp mắt. Mô hình hoạt động cơ bản này hiện diện trong cả chớp mắt do ánh sáng và do không khí gợi lên. Tuy nhiên, việc kiểm tra hồ sơ EMG cho thấy những khác biệt quan trọng trong hoạt động EMG ở các cơ mi đối kháng ngay trong chớp mắt.
Đầu tiên, hoạt động OO khác nhau giữa các lần chớp mắt. EMGOO thể hiện sự bùng nổ hoạt động nhanh chóng để phản ứng với luồng khí (Hình 3A, bản ghi thô). Ngược lại, một kích thích ánh sáng gợi lên sự gia tăng dần dần của EMG và kiểu truyền dẫn đều đặn hơn. Sự khác biệt về tần số của các bùng nổ EMG phản ứng với không khí và ánh sáng phải ảnh hưởng đến vận tốc chuyển động của mí.
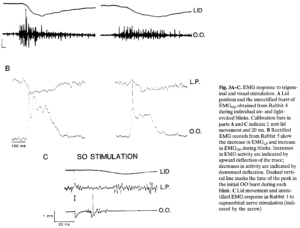
Thứ hai, có sự khác biệt về thời gian hoạt động của các cơ mí mắt tác động qua lại giữa các lần chớp mắt. Để phản ứng với một luồng không khí, EMGLP không hoạt động vào thời điểm xảy ra bùng nổ EMGOO ban đầu (Hình 3B, các bản ghi đã được chỉnh sửa). Do đó, sự co lại ban đầu của OO hầu như không bị LP phản đối. Ngược lại, để phản ứng với một tia sáng, sự giảm EMGLP hầu như không bắt đầu khi xảy ra bùng nổ EMGOO. Do đó, lực co ban đầu của OO phải bị tác động của LP cản trở. Hơn nữa, các bản ghi LP thường cho thấy hoạt động tăng nhẹ so với mức cơ bản ngay trước khi hoạt động giảm liên quan đến chớp mắt do ánh sáng gây ra không rõ ràng trong các lần chớp mắt do không khí gợi lên, điều đó có nghĩa là LP thậm chí còn kháng lại hoạt động của OO nhiều hơn. Sự co lại của các cơ mi đối kháng sẽ làm giảm vận tốc mi mắt hạ xuống so với vận tốc được tìm thấy khi hoạt động của các cơ hạ/đóng mi không bị cản trở. Do đó, sự khác biệt về thời gian hoạt động ở các cơ mí mắt tác động qua lại, xuất hiện trong các lần chớp mắt ở mọi biên độ, góp phần tạo ra sự khác biệt về chuyển động học giữa chớp mắt do ánh sáng và chớp mắt do không khí.
Sự khác biệt giữa các lần chớp mắt có thể phụ thuộc vào phương thức, thay vì chỉ phụ thuộc vào kích thích. Kích thích điện của dây thần kinh sinh ba trên ổ mắt cũng gây ra chớp mắt bằng cách sử dụng con đường sinh ba. Các kích thích tương đối yếu gây ra những chớp mắt nhỏ với đặc điểm nổ EMGOO điển hình của chớp mắt được gợi lên khi kích thích trên ổ mắt (Hiraoka và Shimamura 1977; Kugelberg 1952; Rushworth 1962; Shahani 1970). Độ trễ của phản ứng EMG sớm nhất là khoảng 7 ms kể từ khi kích thích. Sự kích thích điện của dây thần kinh sinh ba trên ổ mắt tạo ra kiểu hoạt động OO và LP theo thời gian giống nhau được tìm thấy với các chớp mắt do luồng không khí gợi lên (Hình 3C, các bản ghi thô). LP ngừng phóng điện trước khi bắt đầu cụm OO và tiếp tục hoạt động sau khi hoàn thành hoạt động OO. Đồ thị vận tốc mi mắt hạ xuống tối đa và thời gian chớp mắt như là một hàm của biên độ chớp mắt mang lại mối quan hệ tương tự đối với các chớp mắt để đáp ứng với kích thích trên ổ mắt hoặc với luồng khí. Cường độ sốc biên độ yếu được sử dụng ở đây tạo ra các chớp mắt không lớn hơn 0,5 mm, do đó chỉ có thể so sánh trong một phạm vi biên độ chớp mắt giới hạn. Tuy nhiên, dữ liệu cho thấy rằng sự khác biệt về thời gian của hoạt động cơ do luồng không khí và ánh sáng tạo ra là do sự kích hoạt thông qua các hệ thống cảm giác khác nhau.
EMG tích hợp tăng lên một cách có hệ thống khi biên độ chớp mắt tăng lên. Các sơ đồ EMG được chuẩn hóa như là một hàm của biên độ của từng lần chớp mắt ở mọi kích thước cho thấy mối quan hệ tuyến tính tương tự đối với cả chớp mắt do không khí và ánh sáng gợi lên. Vì vậy, trong khi tần suất bùng nổ hoạt động OO có thể liên quan đến vận tốc mí, thì tổng lượng hoạt động OO dường như được xác định.
Cường độ kích thích của đặc điểm phản ứng kích thích
Yếu tố chính quyết định cường độ của EMG liên quan đến chớp mắt và cuối cùng là kích thước của chớp mắt là cường độ và thời gian của kích thích gây kích thích chớp mắt. Ví dụ, việc tăng cường độ của một kích thích chặt chẽ trong khi giữ nguyên thời lượng kích thích sẽ dẫn đến EMG lớn hơn (Hình 4)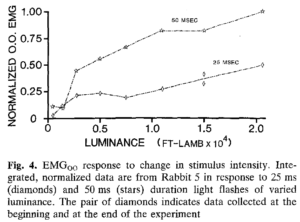 . EMG tăng theo mức tăng cường độ trên một phạm vi rộng các giá trị độ sáng. Với mối tương quan giữa biên độ chớp mắt và hoạt động EMG, không có gì đáng ngạc nhiên khi việc thay thế biên độ chớp mắt cho hoạt động EMG trong sơ đồ này và các sơ đồ tương tự mang lại một con số thực tế giống hệt nhau. Sự gia tăng tương đương về EMGoo và biên độ chớp mắt xảy ra khi tăng cường độ của luồng khí trong thời gian không đổi. Do đó, hoạt động EMGoo và biên độ chớp mắt đều tương quan thuận với cường độ kích thích.
. EMG tăng theo mức tăng cường độ trên một phạm vi rộng các giá trị độ sáng. Với mối tương quan giữa biên độ chớp mắt và hoạt động EMG, không có gì đáng ngạc nhiên khi việc thay thế biên độ chớp mắt cho hoạt động EMG trong sơ đồ này và các sơ đồ tương tự mang lại một con số thực tế giống hệt nhau. Sự gia tăng tương đương về EMGoo và biên độ chớp mắt xảy ra khi tăng cường độ của luồng khí trong thời gian không đổi. Do đó, hoạt động EMGoo và biên độ chớp mắt đều tương quan thuận với cường độ kích thích.
Đặc điểm phản ứng kích thích: thời gian kích thích
Những thay đổi về thời gian kích thích cũng ảnh hưởng một cách có hệ thống đến hoạt động OO. EMG tăng theo cường độ kích thích trong 25 ms và 50 ms nhấp nháy, nhưng kích thích trong thời gian dài hơn sẽ tạo ra phản ứng EMG lớn hơn (Hình 4). Khi cường độ kích thích được giữ không đổi và thời gian kích thích thay đổi, EMG oo và biên độ chớp mắt tăng theo cách tương tự để đáp ứng với các luồng khí hoặc đèn flash thời gian dài hơn (Hình 5A)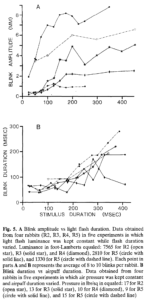 . Đồ thị biên độ chớp mắt so với thời gian kích thích tạo ra một loạt các đường cong vì từng con thỏ khác nhau về phản ứng tương đối với kích thích và vì thỏ được thử nghiệm ở các mức cường độ khác nhau. Tuy nhiên, tất cả các đường cong trong các sơ đồ này đều thể hiện các chức năng có hình dạng tương tự nhau với biên độ nhấp nháy tăng nhanh (hoặc cường độ EMG) đối với các kích thích trong thời gian ngắn hơn và biên độ tăng chậm hơn đối với các kích thích trong thời gian dài hơn. Trên thực tế, phản ứng EMG được chuẩn hóa cho tất cả các đối tượng được vẽ như một hàm của thời gian kích thích mang lại một đường cong duy nhất, tương tự như đường cong sẽ xảy ra nếu dữ liệu trong Hình SA được chuẩn hóa theo biên độ và được biểu thị lại. Do đó, phản ứng chớp mắt với các kích thích không khí hoặc ánh sáng có thời lượng khác nhau sẽ thay đổi một cách đều đặn đối với tất cả các đối tượng.
. Đồ thị biên độ chớp mắt so với thời gian kích thích tạo ra một loạt các đường cong vì từng con thỏ khác nhau về phản ứng tương đối với kích thích và vì thỏ được thử nghiệm ở các mức cường độ khác nhau. Tuy nhiên, tất cả các đường cong trong các sơ đồ này đều thể hiện các chức năng có hình dạng tương tự nhau với biên độ nhấp nháy tăng nhanh (hoặc cường độ EMG) đối với các kích thích trong thời gian ngắn hơn và biên độ tăng chậm hơn đối với các kích thích trong thời gian dài hơn. Trên thực tế, phản ứng EMG được chuẩn hóa cho tất cả các đối tượng được vẽ như một hàm của thời gian kích thích mang lại một đường cong duy nhất, tương tự như đường cong sẽ xảy ra nếu dữ liệu trong Hình SA được chuẩn hóa theo biên độ và được biểu thị lại. Do đó, phản ứng chớp mắt với các kích thích không khí hoặc ánh sáng có thời lượng khác nhau sẽ thay đổi một cách đều đặn đối với tất cả các đối tượng.
Biên độ chớp mắt như là một hàm của thời gian kích thích không khí hoặc ánh sáng luôn tăng nhanh hơn đối với các kích thích có thời gian tương đối ngắn thuộc cả hai loại so với các kích thích có thời gian dài hơn. Điều này xảy ra ở tất cả các biên độ chớp mắt, kể cả những biên độ nhỏ, điều này cho thấy rằng biên độ ổn định không phải do mí mắt trên gặp mí mắt dưới. Thay vào đó, đây là kết quả của chương trình thần kinh tạo ra chớp mắt.
Vẽ đồ thị thời gian chớp mắt như một hàm của thời gian kích thích như được hiển thị đối với luồng khí trong Hình 5B cung cấp cái nhìn sâu sắc về lý do thay đổi độ dốc. Với thời lượng kích thích ngắn hơn 150 ms, thời gian chớp mắt gần như không đổi trong khoảng 65 ms. Việc tăng thêm thời lượng luồng khí sẽ dẫn đến thời gian chớp mắt tăng mạnh. Thời gian chớp mắt tăng theo cách tương tự về mặt chất lượng, gồm hai phần, với kích thích ánh sáng có thời gian dài hơn, ngoại trừ thời gian chớp mắt là khoảng 120 ms với thời lượng kích thích nhỏ hơn 50 đến 75 ms và tăng mạnh khi thời gian kích thích vượt quá 75 ms. Đối với cả không khí và ánh sáng, các giá trị thời gian chớp mắt giữa các đối tượng đều nhất quán một cách đáng kể mặc dù có sự khác biệt về mức độ cường độ kích thích và sự khác biệt về biên độ của chớp mắt. Vì mối quan hệ giữa thời gian kích thích và thời gian chớp mắt không phụ thuộc vào biên độ chớp mắt đối với thời gian kích thích ngắn, nên không có gì đáng ngạc nhiên khi hầu hết các nhà điều tra chỉ báo cáo mối quan hệ yếu giữa biên độ chớp mắt và thời gian chớp mắt (Evinger et al. 1984; Hung et al. 1978; Kennard và Glaser 1984).
Xem xét tổng thể, dữ liệu cho thấy phản ứng OO và chuyển động mí tương ứng xảy ra theo hai giai đoạn riêng biệt. Khi thời gian kích thích không khí hoặc ánh sáng tăng từ 0 ms lên khoảng 150 ms, thời gian chớp mắt vẫn tương đối ổn định trong khi hoạt động EMG và biên độ chớp mắt tăng nhanh. Sau đó, khi thời gian kích thích tăng thêm, thời gian chớp mắt tăng đáng kể trong khi hoạt động E MGoo và biên độ chớp mắt cuối cùng chỉ tăng nhẹ. Điều này cho thấy rằng việc tạo chớp mắt là một quá trình gồm hai bước. Đầu tiên, để đáp lại kích thích ban đầu, một loạt hoạt động OO có tần suất và thời lượng đặt trước sẽ được lập trình. Miễn là kích thích tương đối ngắn, việc tăng thời lượng kích thích sẽ làm tăng cường độ hoạt động EMG, nhưng không làm tăng thời lượng của hoạt động, khiến cho chớp mắt tăng kích thước. Khi thời lượng kích thích đạt đến độ dài tới hạn, các neuron tạo ra chớp mắt sẽ chuyển chế độ hoạt động của chúng. Thay vì tăng tần suất hoạt động của neuron vận động OO, thời gian hoạt động của OO lại tăng lên. Điều này dẫn đến việc kéo dài thời gian chớp mắt đều đặn trong khi chỉ tăng nhẹ biên độ của chớp mắt.
Sự tương tác như vậy giữa biên độ chớp mắt và thời gian chớp mắt có thể thấy rõ khi thay đổi thời gian kích thích, nhưng không thấy rõ khi thay đổi cường độ kích thích. Khi cường độ không khí hoặc ánh sáng tăng lên, cả biên độ chớp mắt và thời gian chớp mắt đều tăng lên ở tất cả các mức cường độ kích thích được sử dụng. Điều này cho thấy rằng sự thay đổi cường độ kích thích bao gồm một quá trình duy nhất ảnh hưởng đến biên độ và thời gian chớp mắt một cách đồng bộ.
Phản ứng EMG với các kích thích có cường độ và thời gian khác nhau
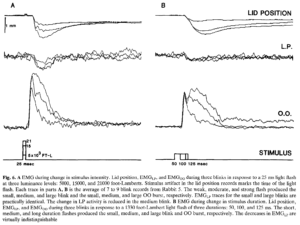
Hoạt động EMG được ghi lại để đáp ứng với các kích thích có cường độ và thời lượng khác nhau phù hợp với phân tích trước đó. Các luồng khí hoặc các tia sáng có cường độ ngày càng tăng có liên quan đến các đợt hoạt động EMG ban đầu lớn hơn và thời gian phân rã dài hơn (Hình 6A). Cũng lưu ý sự thay đổi đều đặn về thời gian đạt đến đỉnh điểm hoạt động EMG và thời gian chuyển động của mí liên quan đến các tia sáng ở ba mức cường độ. Các bản ghi EMG cho thấy hoạt động LP liên quan đến chớp mắt giảm đi kèm theo. Có một xu hướng nhỏ là EMGLP giảm ít hơn với thời gian chớp mắt nhỏ hơn. Vì đèn nháy 25 ms kết thúc trước khi mí bắt đầu di chuyển (lưu ý nhiễu ánh sáng ở dấu vết vị trí mí), những lần nhấp nháy này không thể phụ thuộc vào phản hồi cảm giác. Mỗi lần chớp mắt phải được lập trình riêng trước chuyển động.
Các kích thích không khí hoặc ánh sáng có thời lượng tăng dần nhưng cường độ không đổi tạo ra các đợt hoạt động EMG ngày càng dài hơn và lớn hơn (Hình 6B). Phản ứng EMG đối với kích thích ngắn nhất được mô tả, một tia sáng 50 ms, là một hoạt động bùng nổ đơn lẻ, sắc nét và giảm dần. Để đáp lại kích thích 100 ms, một đợt bùng phát ban đầu tương tự xảy ra, theo sau là đợt hoạt động thứ hai mới xuất hiện. Phản ứng với kích thích 125 ms bắt đầu bằng một đợt hoạt động cũng gần giống với đợt bùng phát ban đầu liên quan đến các lần nhấp nháy 50 và 100 ms, nhưng tiếp tục với đợt hoạt động thứ hai lớn hơn cả về cường độ và thời lượng so với đợt hoạt động liên quan đến kích thích 100 ms. Biên độ của xung ban đầu phản ánh cường độ của kích thích phát ra. Bùng nổ sớm này, đại diện cho toàn bộ phản ứng chớp mắt với các kích thích rất ngắn và chỉ là phản ứng sớm với các kích thích trong thời gian dài hơn, là một kích thích không đổi và trong thời gian dài không còn đánh dấu thời điểm đạt đến đỉnh điểm tổng thể của bùng nổ OO. Thay vào đó, đỉnh điểm của hoạt động EMG xảy ra vào những thời điểm ngày càng muộn hơn trong bùng phát thứ cấp. Bùng nổ thứ cấp chỉ bắt đầu phân rã khi quá trình kích thích hoàn tất. Vì phản ứng EMG phản ánh khoảng thời gian của kích thích nên các neuron vận động chớp mắt phải phụ thuộc vào thông tin cảm giác hiện tại để duy trì chớp mắt. Do đó, một đường dẫn chớp mắt bắt đầu hoạt động của neuron vận động OO dựa trên cường độ kích thích khi kích thích bắt đầu. Con đường thứ hai dài hơn hoặc chậm hơn sau đó duy trì hoạt động OO bằng cách sử dụng thông tin kích thích hiện tại. Sự suy giảm hoạt động EMGLP là tương tự đối với tất cả các lần chớp mắt và gợi nhớ đến phản ứng với các kích thích có cường độ khác nhau.
Thảo luận
Nháy mắt có nhiều hình thức khác nhau
Chuyển động học của mi và mô hình hoạt động EMG ở thỏ cho thấy hai loại phản xạ chớp mắt riêng biệt. Nháy mắt do không khí tạo ra đạt được vận tốc tối đa cao hơn và cần ít thời gian hoàn thành hơn so với chớp mắt do ánh sáng tạo ra có biên độ bằng nhau. Hai yếu tố dường như tạo nên sự khác biệt về động học này là tần số ban đầu của hoạt động OO và thời gian hoạt động ở các cơ mí mắt tác động qua lại. EMG do không khí gợi lên ban đầu nhanh hơn EMG được gợi lên bởi ánh sáng và LP thư giãn trước khi kích hoạt OO trong các lần nháy mắt do không khí gợi lên, nhưng vẫn hoạt động cho đến sau khi bùng nổ OO bắt đầu trong các nháy mắt do ánh sáng gợi lên. Do mối quan hệ giữa EMG tích hợp và biên độ chớp mắt là tương tự nhau đối với cả chớp mắt do ánh sáng và do không khí tạo ra, nên tổng lượng hoạt động OO đi kèm với các chớp mắt do không khí và do ánh sáng tạo ra có biên độ bằng nhau là tương đương nhau. Điều này ngụ ý rằng một neuron vận động OO riêng lẻ phát ra tần số truyền dẫn cao hơn trong một chớp mắt do không khí tạo ra. Tuy nhiên, vì chớp mắt do ánh sáng tạo ra đòi hỏi thời gian phóng điện dài hơn nên neuron vận động OO có thể tạo ra cùng một điện thế hoạt động đối với chớp mắt do không khí và do ánh sáng tạo ra có biên độ bằng nhau. Các neuron bùng nổ dẫn trung bình, các tế bào hướng tâm tiền vận động của các neuron vận động ngoại nhãn tạo ra các chuyển động giật (saccadic) của mắt, biểu hiện một mô hình hoạt động tương tự. Trong một lần giật, các neuron thường không hoạt động này tạo ra một đợt hoạt động tần số cao trong đó tần số truyền dẫn tương quan với vận tốc giật tối đa và số lượng nhọn sóng trong lần bùng nổ tương quan với biên độ giật (Keller 1974; King và Fuchs 1979). Vì vậy, neuron tạo ra chớp mắt có thể có các đặc tính tương tự như neuron tạo ra giật mắt.
Sự khác biệt giữa chớp mắt do không khí và ánh sáng kích thích có nghĩa là các neuron tiền vận động điều chỉnh hoạt động LP và OO là riêng biệt hoặc chuỗi sự kiện trong một tập hợp các tế bào tiền thần kinh vận động chung phải khác nhau để đáp ứng với hai loại kích thích. Dữ liệu trên ổ mắt cho thấy rằng, bất kể cơ chế nào, sự khác biệt là ở phương thức, hơn là theo kích thích cụ thể, phân loại theo phương thức có thể là cách hữu ích nhất để nhóm các dạng chớp mắt phản xạ. Chớp mắt của con người tương tự về mặt chuyển động học với chớp mắt của thỏ (Evinger et a1. 1984), bao gồm các con đường ly tâm và hướng tâm giống nhau, và dữ liệu sơ bộ (Doane 1980; Manning và Riggs, dữ liệu chưa được công bố) cho thấy rằng các dạng chớp mắt khác nhau có thể cũng có mặt ở người.
Các kích thích sinh ba và thị giác gợi lên sự di chuyển xuống của mí mắt bằng cách sử dụng cùng một nhóm cơ, nhưng các chuyển động rất khác nhau. Trong nghiên cứu về điều khiển vận động, đây là một ví dụ về “sức mạnh tổng hợp” hoặc “hình thức” chuyển động của cơ. Khái niệm này nảy sinh từ việc phân tích các chuyển động đa hình thức trong đó hệ thống thần kinh trung ương dường như đơn giản hóa việc kiểm soát chuyển động bằng cách tự giới hạn ở một số kiểu co cơ có thể có khả năng đưa chi đến một điểm cuối nhất định. Việc tạo ra các dạng chuyển động riêng biệt có thể được thực hiện bằng cách thay đổi thời gian co cơ tương đối. Hình thức cụ thể của chuyển động được gợi ra phụ thuộc vào kích thích. Ví dụ, rùa gãi cơ thể bằng ba cử động chi sau riêng biệt (Mortin và cộng sự 1985; Robertson và cộng sự 1985), tùy thuộc vào vị trí của chất kích thích. Các hình thức gãi khác nhau là do sự thay đổi về thời gian co cơ của chi. Tương tự như vậy, để khôi phục lại tư thế thẳng đứng cân bằng, đối tượng con người gợi lên một trong hai chuyển động tạo thành các dạng khác nhau, sức mạnh tổng hợp của mắt cá chân hoặc sức mạnh tổng hợp của hông (Nashner và McCollum 1985), tùy thuộc vào cơ sở hỗ trợ. Một lần nữa, sự thay đổi về thời gian co cơ trong cùng một nhóm cơ làm nền tảng cho hai sự phối hợp này.
Nháy mắt thể hiện các hình thức hoặc sự phối hợp theo cách tương tự. Các chuyển động khác biệt về mặt chuyển động học đạt được cùng một mục đích là hạ mi mắt xuống, nhưng chúng khác nhau về thời điểm co cơ mí mắt, điều này phụ thuộc vào bản chất của kích thích. Sự đơn giản tương đối của tổ chức thần kinh dựa trên chớp mắt cung cấp một hệ thống vận động lý tưởng để phân tích khái niệm về sự phối hợp cơ ở cấp độ tế bào.
Kiểm soát kích thích chớp mắt
Mặc dù có nhiều hình thức chớp mắt khác nhau được gợi lên thông qua các hướng tâm thị giác và sinh ba, nhưng tất cả các chớp mắt đều có chung các đặc điểm cơ bản. Biên độ chớp mắt và yếu tố quyết định chính của nó, hoạt động OO, tương quan thuận với cường độ kích thích, một đặc điểm chung của phản xạ (ví dụ, Sherrington 1948). Quan trọng hơn, hai hình thức điều khiển thần kinh dường như hoạt động trong các chớp mắt phản xạ. Thành phần sớm nhất của chuyển động mí được lập trình trước chuyển động. Bất kể khoảng thời gian cuối cùng của kích thích là bao lâu, chuyển động ban đầu của mi mắt là như nhau đối với kích thích có cường độ không đổi. Nếu kích thích kết thúc trước khi bắt đầu chớp mắt để chuyển động của mi mắt không làm thay đổi đầu vào cảm giác thì chớp mắt vẫn bắt đầu theo cách tương tự nhưng kết thúc sau một thời gian đã định. Trong trường hợp này, các neuron sản xuất chớp mắt sẽ lập trình toàn bộ quá trình chớp mắt, từ đầu đến cuối, trước khi bắt đầu chuyển động. Với các kích thích có thời lượng ngày càng tăng, thời gian hoạt động OO sẽ kéo dài hơn. Điều này ngụ ý rằng sau phần chớp mắt sớm được lập trình trước, việc chớp mắt sẽ tự động kết thúc sẽ không xảy ra. Thay vào đó, việc khép mí nằm dưới sự kiểm soát kích thích. Cường độ và thời lượng bùng nổ OO cuối cùng phản ánh cường độ và thời lượng kích thích.
Kiểu phản ứng này, bắt đầu bằng thành phần không thay đổi, được đặt trước và kết thúc bằng thành phần nhạy cảm hơn, có thể điều chỉnh được, được tìm thấy trong các phản xạ khác, nhiều trong số đó như chớp mắt (Evinger và Manning 1985) cho thấy khả năng học tập thích ứng. Ví dụ, phản xạ tiền đình (VOR) thể hiện một tổ chức gồm hai phần tương tự (Lisberger 1984), trong đó kích thích tiền đình nhanh chóng gợi lên một chuyển động bù trừ ban đầu của mắt, sau đó là thành phần thứ cấp của chuyển động 5 ms sau đó. Việc thay đổi độ lợi của VOR sẽ làm thay đổi thành phần thứ hai một cách có hệ thống, nhưng không làm thay đổi thành phần thứ nhất. Dữ liệu như vậy cho thấy thành phần thứ cấp của chớp mắt có thể là phần phản xạ chớp mắt có thể được sửa đổi thông qua sự kiểm soát trung tâm hơn.
Nguồn:



Đăng nhập để bình luận.